
Stinging Nettle: More Than a Weed, More Than a Cure
Food, fibre, medicine, butterfly habitat, soil signal, and regenerative ally, the forgotten power of one of the world’s most misunderstood plants.

Urtica dioica — Stinging Nettle
A Deeper Cut: The Nettle Monograph, Rebuilt
Last year I posted a Living Plant Wisdom Profile on stinging nettle. It was a good start. It deepened my appreciation for a plant I already loved. But the question underneath had been there from the beginning: how do you compile the most complete monograph on a plant and keep it useful to people who actually want to work with nature?
After more revisions than I care to count, I think I’ve found the shape. Nettle was the first plant I profiled here, so it earned the first full pass under the new framework. No paywall. Read it end to end, sit with the parts that matter to you, come back to it as the seasons turn. Let me know what you think?
Fair warning: the new profile is longer, denser, slower. It’s built for land stewards working at any scale, from a single hedgerow patch to a managed hectare, and it trades easy reading for usefulness in the field. What I’m after isn’t another reference document. It’s a way of meeting a plant, clearly enough, honestly enough, that the next time you see it growing, you see something you didn’t see before.
That shift, as small as it sounds, is what changes how a steward works.
This 21-section Ontology is a framework for examining any plant worth knowing through the same disciplined lens: botany, ecology, ethnobotany, phytochemistry, folklore, cross-cultural convergence, and the honest speculative edges where the record runs thin.
The result isn’t just more thorough, it’s clearer in ways the original couldn’t be. Some of what I wrote last year was right but partial. Some of it I now see differently. That’s what a real lens does: it doesn’t just add detail, it adjusts. It shows you where you were squinting.
The first was species clarity. Most of what you read about “stinging nettle” in North American herbals, Indigenous ethnobotany sources, and modern supplement marketing is about Urtica gracilis, the native North American plant, diploid, often monoecious, not Urtica dioica, the Eurasian tetraploid. Kew restored gracilis to species rank in 2023–2024. Most field guides haven’t caught up. Most herbal writing still conflates them. The new profile names the plant each source is actually talking about.
The second was the "dynamic accumulator" myth. I had written it, repeated it, believed it: that nettle "mines minerals from deep soil." After more digging, the claim traces to two grey-literature sources in the early 1980s, neither presenting experimental evidence. The plant's rooting architecture doesn't support it. The foliar mineral content is real; the deep-mining story is not validated.
The third was cross-cultural convergence. When five or six unrelated traditions, Dioscorides, Nlaka’pamux, Ibn Sīnā, Tibetan zwa-ma, Slavic Maundy Thursday, Roman urtication, all point to the same function in the same plant, that’s evidence. Not proof. But evidence strong enough to map onto the chemistry and ask: what compound class is the shared thread? The new profile does this work for six convergences: hemostatic, counter-irritant for rheumatic pain, spring mineral tonic, diuretic, BPH-specific root use, and bast fibre. Each carries a research frontier hypothesis that would translate traditional knowledge into testable modern pharmacology.
The fourth was honesty about gaps. The new profile flags over twenty specific points where the evidence runs out, where a claim is widely repeated but never tested, where a chemistry study has been done on European nettle but not on North American gracilis, where a traditional use has never been clinically verified. Silence is data. Pretending otherwise is what makes herbal writing rot.
Below is the full table of contents. Take what’s useful. Come back to the sections that matter to your practice.
A note on what comes next: profiles like this take a large amount of time, researching, reading, cross-referencing, and putting this together in a way that makes sense to as many people as possible. Going forward, the first four sections of each Living Plant Wisdom Profile will stay free, enough to meet the plant, enough to know whether the rest is for you. The deeper material, the convergence work, the chemistry, the stewardship sections, will sit behind a paywall for paid subscribers. That’s how this work stays sustainable, and how it stays the kind of work it needs to be: slow, careful, accountable to the plant rather than to the algorithm.
This one’s the gift. I hope you find a friend in it.

Table of Contents
Phase I — The Plant in Its World
The ecology, the identity, the relationships that existed before any human wrote anything down.
1. Plant Identity Snapshot — taxonomy, range, and the gracilis split explained in plain English.
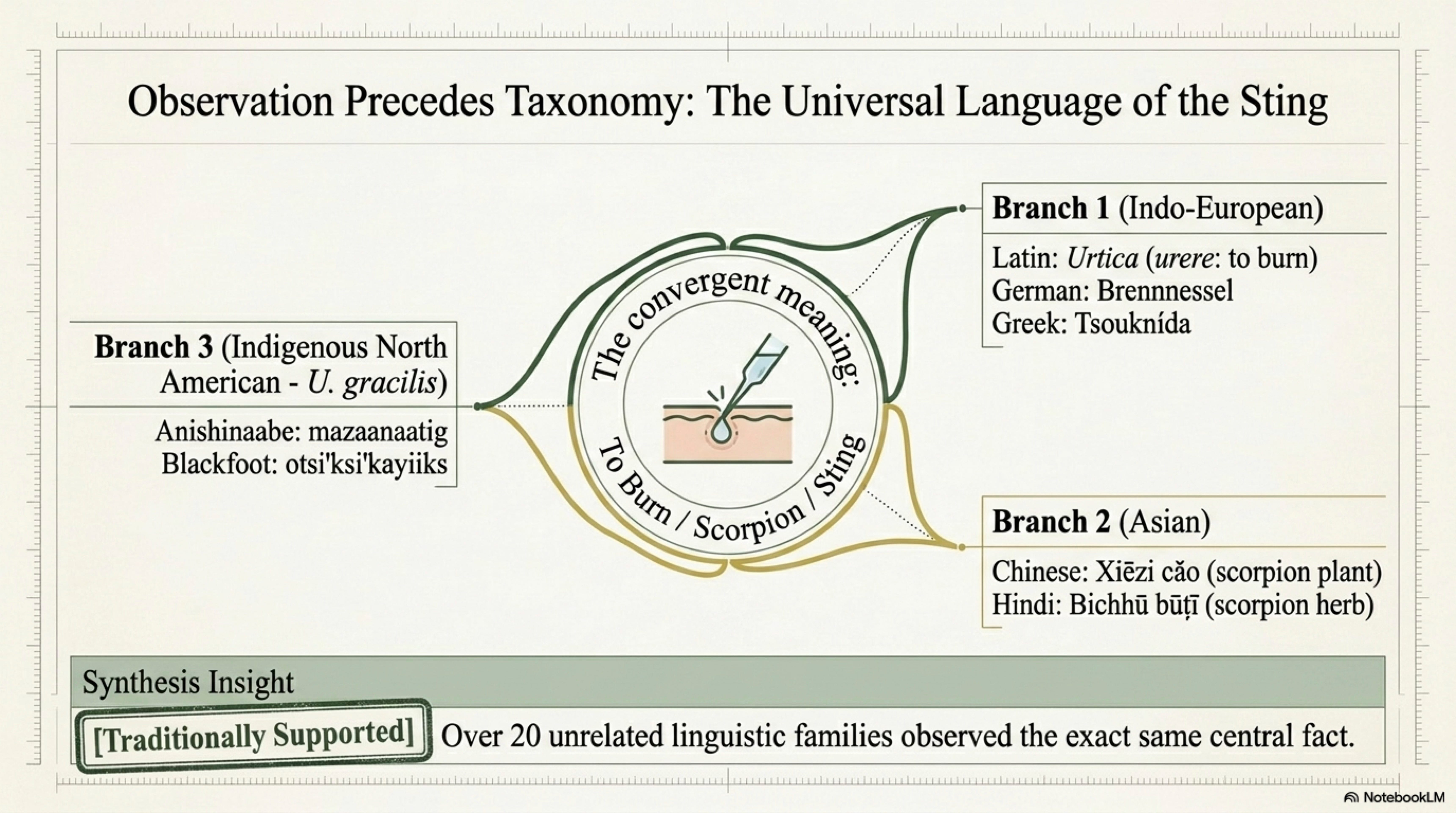
2. Names, Language, and Lineage — 20+ languages surveyed; the convergent “burning” etymology across unrelated linguistic families; Indigenous names attributed to specific nations with their documenting sources.
3. Identification and Look-Alikes — wood nettle, false nettle, dead-nettle, horse nettle, clearweed; the three-second field check.
4. Botanical Character and Life Cycle — rhizome intelligence, seed bank persistence, clonal longevity, and why a nettle patch is less a crowd than a family.
5. Ecological Intelligence — soil, water, community, pollinators, ecosystem function, and the indicator-value chart. The dynamic-accumulator myth addressed directly.
6. Animal Interactions and Ethology — the mammal-avoidance/insect-specialization paradox, nymphalid butterfly dependency, zoopharmacognosy honestly evaluated.
7. Climate Resilience and Adaptation — why nitrogen deposition matters more than temperature for nettle’s range shifts, and what that means for regenerative practice going forward.
8. Phenology and Working Calendar — harvest windows tied to sensory cues, not just calendar dates. “Nettle time” as a two-to-three-week annual event per patch.
9. History, Folklore, and Cultural Memory — Lusehøj Bronze Age textile (imported across Europe), the Nine Herbs Charm, Andersen’s Wild Swans, the encoded agronomy of “nettle in, dock out.”
10. Traditional Ecological Knowledge and Stewardship — CARE principles applied; seventeen Indigenous nations cited with source attribution; attributional ethics throughout.
Phase II — The Plant in Human and Animal Hands
What we’ve done with it. What the traditions say. What the chemistry confirms, and where they diverge.
11. Food, Medicine, and Human Use Traditions — Western herbal, TCM, Ayurveda (and its absence), Unani, Tibetan, Indigenous North American, Andean, Himalayan, with cross-cultural synthesis identifying six convergent uses.
12. Chemistry, Nutrition, and Functional Compounds — complete nutritional profile, phytochemistry by compound class, the UDA lectin antiviral story (HIV, CMV, SARS-CoV), chemistry-tradition convergence screen mapping six cultural claims onto specific compound classes.
13. Safety and Responsible Use — oxalate, drug interactions, pregnancy (where tradition and modern caution diverge), heavy metal accumulation, sourcing ethics.
14. Regenerative Agriculture and Land Applications — purin d’ortie recipe and microbiology, KNF FPJ adaptation (nettle-specific), biodynamic preparation 504 with honest evidence review rather than either dismissal or boosterism.
15. Homestead and Material Uses — bast fibre from Bronze Age to STING project; Pacific Northwest whaling-line tradition; dye; the notable absence of nettle as a smudge herb.
16. Harvest, Processing, and Preservation — sensory quality indicators (smell, taste, touch, colour, sound) for field practitioners. What your hands and nose tell you the lab confirms.
17. Economics and Practical Value — patch-scale case-study math, replacement value for farm inputs, and the resilience-economics argument for why marginal-land plants matter in a volatile future.
18. Legal, Regulatory, and Access Notes — the full purin d’ortie regulatory saga from French AMM requirement through the 2017 EU basic-substance approval. A paradigm case for traditional practice colliding with modern regulation — and winning.
Phase III — The Honest Edges
Where the evidence runs out, where the metaphors begin, and where the questions worth asking still live.
19. Research Frontiers and Open Questions — 22 specific gaps flagged, from North American gracilis phytochemistry (nobody’s actually done it) to UDA lectin pandemic relevance to the pregnancy-safety evidence gap.
20. Speculative, Symbolic, and Relational Layer — every claim labeled as Metaphor, Belief, or Frontier Hypothesis. Signature readings without the woo. A discipline for talking about what a plant teaches without pretending it’s what a plant proves.
21. Sources, Confidence, and Citation Architecture — five-tier confidence tagging (Well-documented → Traditionally supported → Emerging → Anecdotal → Speculative, with Gap flags throughout), 200+ cited sources, living-document notes for future revisions.
What’s genuinely new
For readers who had the first version:
Species clarity throughout — the plant you thought you knew might be a different one
Inline citations on every weight-bearing claim
Confidence tags distinguishing Well-documented from Emerging from Speculative
Honest gap-flagging where the record is silent
Cross-cultural convergence methodology — six claims validated across three or more unrelated traditions
Chemistry-tradition mapping — which compound class carries which cross-cultural claim
Indigenous attribution at the level of specific nations and documenting ethnobotanists, not flattened “Native American”
The purin d’ortie regulatory saga as a full case study in traditional-practice vs. modern-regulation
MBFH labeling in the speculative section — the discipline that lets us talk about what a plant teaches without pretending it’s what a plant proves
The dynamic-accumulator claim addressed directly rather than repeated
The monograph is a living document. Corrections welcome. Gaps are listed for a reason, if you have peer-reviewed work that closes one of them, I want to hear about it.

Phase I — The Plant in Its World
1. Plant Identity Snapshot
Common names: stinging nettle, common nettle, burn nettle, burn hazel, Brennnessel, grande ortie, ortica, крапива, tsouknída, 荨麻 (xúnmá), sisnu.
Latin binomial: Urtica dioica L. (1753, Species Plantarum 2:983)
Family: Urticaceae (the nettle family — ~53 genera, ~2,600 species worldwide)
Native range (L. s.s.): Europe, western and central Asia, North Africa, Macaronesia [POWO 2026; Taylor 2009].
Introduced and widely naturalized: North America, South America (temperate), Australasia, southern Africa [CABI 2023].
Current regional status: common to superabundant on nitrogen-enriched ground across the Holarctic; not formally invasive in most jurisdictions because it is also native across much of the range where it is abundant; weedy but ecologically native in Britain, continental Europe, western Russia.
Synonyms and sister taxa: U. dioica subsp. dioica (the tetraploid, strictly dioecious Eurasian type, 2n=52); subsp. holosericea (western North America); subsp. gansuensis and subsp. afghanica (Asian); and, crucially for this profile, Urtica gracilis Aiton, the diploid (2n=26), often monoecious North American native that POWO now accepts as a distinct species and that earlier floras lumped under U. dioica [POWO 2026; GRIN 2024; Boufford 1997; Bassett et al. 1974]. [Well-documented]
One-sentence thesis. Urtica dioica is the plant that marks the places where humans have lived, the middens, byres, compost-piles, riverbanks, and disturbed woodland edges where nitrogen and phosphorus have accumulated, and it responds to that ground by building protein, pigment, fiber, and pharmacy at rates few other temperate herbs can match.
Relationship thesis. Nettle keeps no secrets. The sting is a promise: respect the hand that approaches, and the plant will offer back more than it takes. Nowhere on earth have humans lived near nettle without learning to handle it; nowhere has the lesson failed to pay forward in food, in cloth, in medicine, in soil.

2. Names, Language, and Lineage
2.1 Scientific identity
Accepted name: Urtica dioica L., Species Plantarum 2:983 (1753). [Well-documented]
Taxonomic history. Linnaeus established the binomial in 1753, selecting “dioica”, “two-housed”, to mark the separation of male and female flowers onto separate plants. Aiton, in Hortus Kewensis (1789), described U. gracilis from North American material. Nineteenth- and twentieth-century treatments lumped gracilis as a subspecies or variety of U. dioica; POWO’s 2023–2024 revisions restored species rank [POWO 2026]. [Well-documented]
Key subtaxa (historically recognized under U. dioica s.l.):
subsp. dioica, Eurasian type, tetraploid, strictly dioecious
subsp. gracilis (Aiton) Selander → now U. gracilis Aiton, diploid, often monoecious, North American native
subsp. holosericea (Nutt.) Thorne, western North America, now often treated under U. gracilis
subsp. gansuensis C.J. Chen, northwestern China
subsp. afghanica Chrtek, Afghanistan and adjacent mountains [POWO 2026; Flora of China Vol. 5; GRIN 2024]. [Well-documented]
Chromosome number. Subsp. dioica: 2n=52 (tetraploid, base x=13). U. gracilis s.s.: 2n=26 (diploid). subsp. holosericea: 2n=26 or 52, population-dependent [GRIN 2024; Bassett et al. 1974]. [Traditionally supported, consistent across multiple cytological studies but no post-split synthesis.]
Sister species worth naming:
U. urens L., small nettle, dwarf nettle. Annual, monoecious, nitrogen-demanding, common in gardens and row-crop fields. Smaller in every dimension.
U. pilulifera L., Roman nettle. Annual, monoecious, spherical female inflorescences.
U. ferox G. Forst., New Zealand tree nettle (ongaonga). The only nettle known to have caused human fatality (one documented case, 1961), and occasional dog and horse deaths [Connor 1977]. [Well-documented]
Girardinia diversifolia, Himalayan allo nettle. Often confused with U. dioica in the Himalayan fiber literature; the two are distinct genera but share range and use. [Well-documented]
2.2 Names across cultures
Indo-European (European and classical):
Latin: Urtica, from urere, “to burn” [Pliny NH XXII.13; Virgil Georgics III.314]
Ancient Greek: ἀκαλήφη (akalēphē), κνίδη (knidē), the latter from the root “to sting,” surviving in botanical Cnidium, cnidaria (jellyfish) [Dioscorides IV.93; Theophrastus HP 7.7]
Old English: netele; and the ceremonial name wergulu, a word that appears uniquely in the Nine Herbs Charm of the Lacnunga (Harley MS 585, 10th–11th c.) and whose philological root is still debated [Pettit 2001; Cameron 1993]
German: Brennnessel, Große Brennnessel, Donnernessel (”thunder-nettle,” against lightning) [Marzell IV]
Russian: крапива (krapíva), krapíva dvudomnaya (”two-housed”) [Annenkov 1878]
Modern Greek: τσουκνίδα (tsouknída) [Heldreich 1862]
Welsh: danadl poethion (”hot nettles”); Irish Gaelic neantóg; Scottish Gaelic feanntag, deanntag
Romance: French grande ortie, Italian ortica comune, Spanish ortiga mayor, Portuguese urtiga-maior
West Asian and Middle Eastern:
Arabic: قُرَّاص (qurrāṣ); Maghrebi أنجرة (anjura) [Ibn al-Bayṭār, Al-Jāmiʿ]
Persian: گزنه (gazneh) [Schlimmer 1874]
Turkish: ısırgan otu (”biting plant”) [Baytop 1999]
Hebrew: סִרְפָּד (sirpad), appears in Isaiah 55:13, Hosea 9:6, poetically identified with nettle though the exact species in the biblical landscape is contested [Feliks, Plant World of the Bible]
South and Central Asian:
Sanskrit: vṛścikālī (वृश्चिकाली, “scorpion-like”) appears in classical materia medica but is more reliably identified with Tragia involucrata (a stinging Euphorbiaceae) than with Urtica dioica [Nadkarni 1908; Kirtikar & Basu III]. [Traditionally supported for the word; species attribution uncertain.]
Hindi: बिच्छू बूटी (bichhū būṭī, “scorpion herb”), kandali [Watt, Dict. Econ. Products]
Nepali: sisnu (सिस्नु), often covers U. dioica and Girardinia diversifolia together in Himalayan use [Manandhar 2002]
Tibetan: ཟྭ་མ (zwa ma); སྦྲུལ་ཤིང (sbrul shing, “snake wood”) [Pasang Yonten Arya 1998]
East Asian:
Chinese: 蕁麻 / 荨麻 (xúnmá); also 蜇人草 zhērén cǎo (”stinging-people plant”) and 蠍子草 xiēzi cǎo (”scorpion plant”) [Zhonghua Bencao 1999 vol. 2]
Japanese: イラクサ (irakusa, 刺草, “thorn plant”)
Korean: 쐐기풀 (ssaegipul)
Mongolian: хорголзгоно (khorgolzgono) [Ligaa 1996]
Indigenous North American, U. gracilis lineage. Each name is attributed to the nation whose knowledge keepers recorded it with the cited ethnobotanist. These names belong to those communities; they are cited here with the same care a practitioner would give a quoted line.
Anishinaabemowin (Ojibwe): mazaanaatig, mazaana [Densmore 1928]
Plains Cree: masān, maskosiwi-masān [Leighton 1985]
Blackfoot: otsi’ksi’kayiiks [Hellson 1974]
Cherokee: ᎤᎩᎬᎯᏓ (ugigvhida) [Hamel & Chiltoskey 1975]
Menominee: masā́nask [Smith 1923]
Lakota: čhaŋȟlóǧaŋ ičáȟpe [Rogers 1980]
Halkomelem (Central Coast Salish): ts’ítx̱ʷəɬp [Turner & Bell 1971]
Kwak’wala (Kwakwaka’wakw): ǥałǥadi̓ [Turner & Bell 1973]
Nuu-chah-nulth: ḥiḥinkʷałaqƛ [Turner & Efrat 1982]
Diné (Navajo): gah ałchʼį́ʼ ditłʼoʼí [Wyman & Harris 1941]
2.3 Meaning of names
The names agree on one thing: this plant burns. Urtica and krapíva and Brennnessel and xiēzi cǎo and bichhū būṭī all derive from verbs of stinging, scorching, scorpion-bite. Twenty-odd unrelated languages have looked at the same herb and chosen the same central fact to carry in the name [Traditionally supported; cross-linguistic survey per §2.2 evidence file]. That convergence alone, with no shared linguistic root, is one of the cleaner demonstrations that observation precedes taxonomy. The sting is the first thing a human notices; the sting is what the name preserves.
The second thing the names reveal is place. Donnernessel, thunder-nettle, kept in Alpine windowsills to catch lightning. Sbrul shing, snake wood, Tibetan shorthand for the coiled quality of rhizomes. Mazaanaatig, Anishinaabe for something close to “basket-plant,” the cordage recognized in the naming. The scientific epithet dioica, two-housed, encoded the botanical observation that male and female flowers live on separate stems, which is true of the European tetraploid but not always of the North American diploid [Bassett et al. 1974]. The names are sharper than the taxonomy because the naming was older than the microscope.
What the names don’t say, the silences say. There is no name for U. dioica in the classical Sanskrit materia medica of the Indian heartland, the plant is a Himalayan borderland herb in India, not a plains plant, and the southern schools of Ayurveda simply did not develop a monograph on it [Chopra et al. 1956; Warrier et al. 1994]. Silence is data. The northern Himalayan Amchi traditions have a full working pharmacopoeia of zwa-ma [Pasang Yonten Arya 1998]; the shastra of Caraka does not. Knowing where the tradition runs out matters as much as knowing where it runs deep.
3. Identification and Look-Alikes
3.1 Field identification
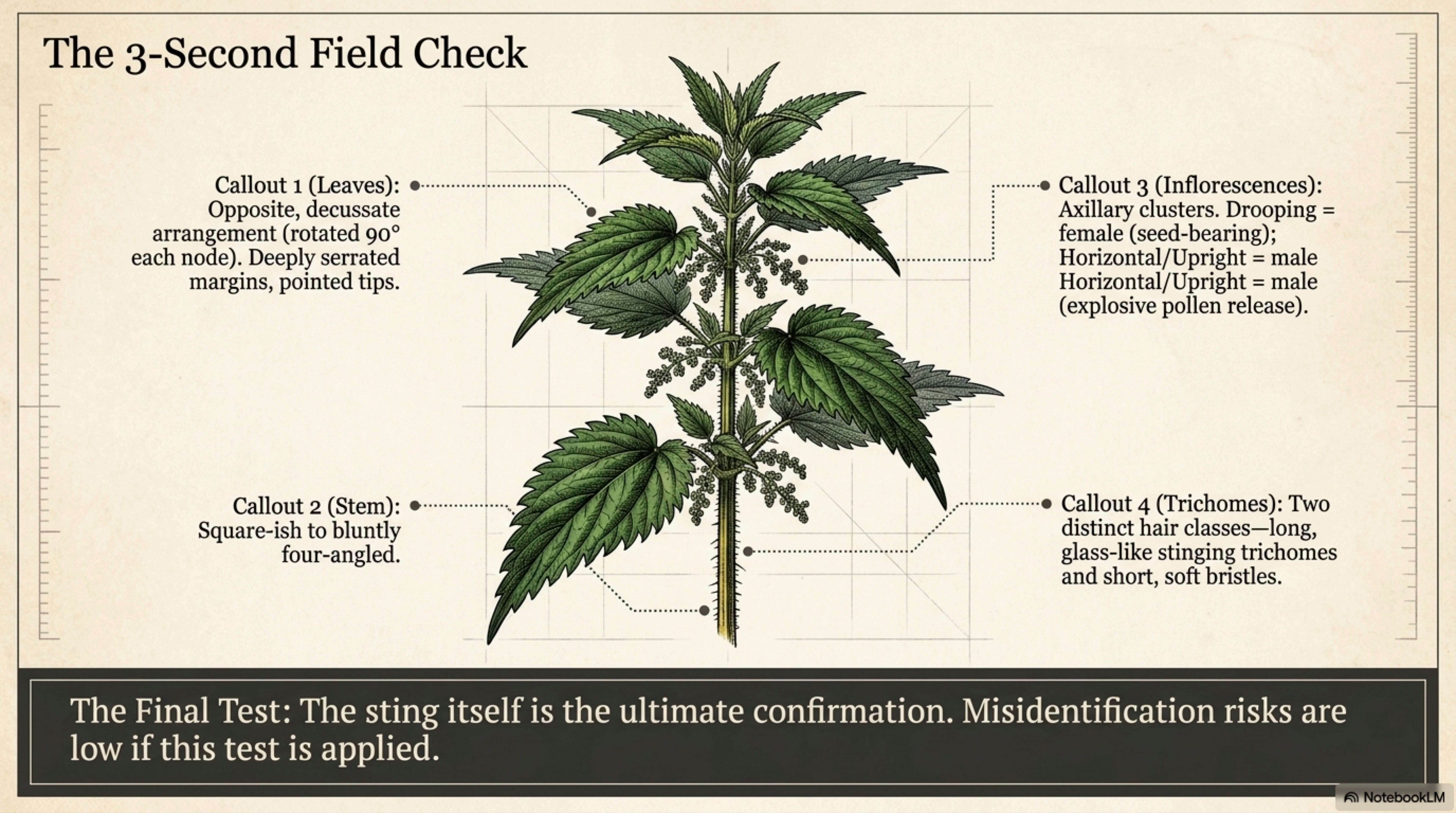
Growth habit. Upright, unbranched or sparsely branched perennial herb, 50–200 cm in mature patches, rising from a dense network of yellow rhizomes that spread horizontally at 5–15 cm depth [Taylor 2009]. Where soil is fertile and moist, nettle forms pure stands, a knee- to shoulder-high green wall, often several meters across, sometimes a hectare. The stands are clonal: what looks like a crowd is often a few families. [Well-documented]
Stem. Square-ish to bluntly four-angled, erect, covered in two sizes of hair, long stinging trichomes and shorter non-stinging bristles. Young stems green; older stems sometimes tinged purple at nodes. Hollow in the lower reaches on vigorous plants [Taylor 2009]. Run a finger up a stem and the direction of the needles tells you: swept toward the tip, like scales.
Leaves. Opposite, decussate (successive pairs rotated 90° from each other), ovate to ovate-lanceolate, 4–15 cm long, with deeply serrate margins and a sharply pointed tip. Leaf surface bears the same two trichome classes as the stem. Young spring leaves may be almost black-green and bronze at the tip; midsummer leaves settle to a matte, slightly glaucous green [Boufford 1997; Taylor 2009]. The undersurface is paler; the three main veins arch from near the base.
Flowers. Small, greenish, wind-pollinated, borne in catkin-like axillary inflorescences 3–10 cm long. Male flowers held upward or horizontal; female flowers typically held downward, denser, more branched. On a hot windless June morning in a mature stand, a sharp knock against a male inflorescence will release a visible pollen cloud, the explosive stamen dehiscence is one of the small theaters of the plant world [Taylor 2009]. [Well-documented]
Dioica: male and female flowers on separate plants (strict in Eurasian populations). Gracilis: frequently monoecious in North America, both sexes on the same plant, sometimes in the same inflorescence [Bassett et al. 1974; Boufford 1997]. This is the most reliable field distinction between the two lineages.
Seed (achene). Small (~1–1.5 mm), flattened, olive-brown, hidden among persistent perianth segments in dense pendulous female inflorescences. A single mature female stem can bear thousands [Taylor 2009].
Root. Rhizomatous. Yellow cortex, bright when freshly dug, with fibrous roots branching off the rhizome at short intervals. Rhizomes can persist at least a decade in undisturbed patches; individual ramets shorter-lived [Taylor 2009]. On close-in examination the rhizome smells faintly of turnip and damp humus.
Smell. Crushed fresh leaf: clean, green, slightly iodine-like, with an undertone often described as “algal” or “marine.” Dried leaf: more hay-like, with a distinct mineral-sweet note from chlorophyll degradation products.
Texture. Fresh young leaf: soft, almost velvety on the upper surface when the trichomes have not yet calcified. Fresh mature leaf: papery, with the trichomes fully stiff. Stem past flowering: fibrous, beginning to “ret”, the signal that the bast fiber is developing.
Habitat clues. If the patch is thick, tall, uniform green, ankle-knee-shoulder tall, growing in a river terrace, a hedge base, a compost heap edge, a disused garden corner, a cow-camp, the edge of a chicken run, or a place where sheep have sheltered in a gap for seasons on end, it is almost certainly nettle. The plant is a living receipt for nitrogen history.
Key field marks (three-second check). (1) Opposite leaves with deep serrations and pointed tips. (2) Stem and leaves bearing two sizes of hair, one class unmistakably a stinging needle when the light catches it. (3) Square-ish stem. (4) Inflorescences in the leaf axils, drooping when female, horizontal when male. (5) The sting itself, when sleeved skin accidentally brushes the plant, the final and unmistakable confirmation.
3.2 Look-alikes
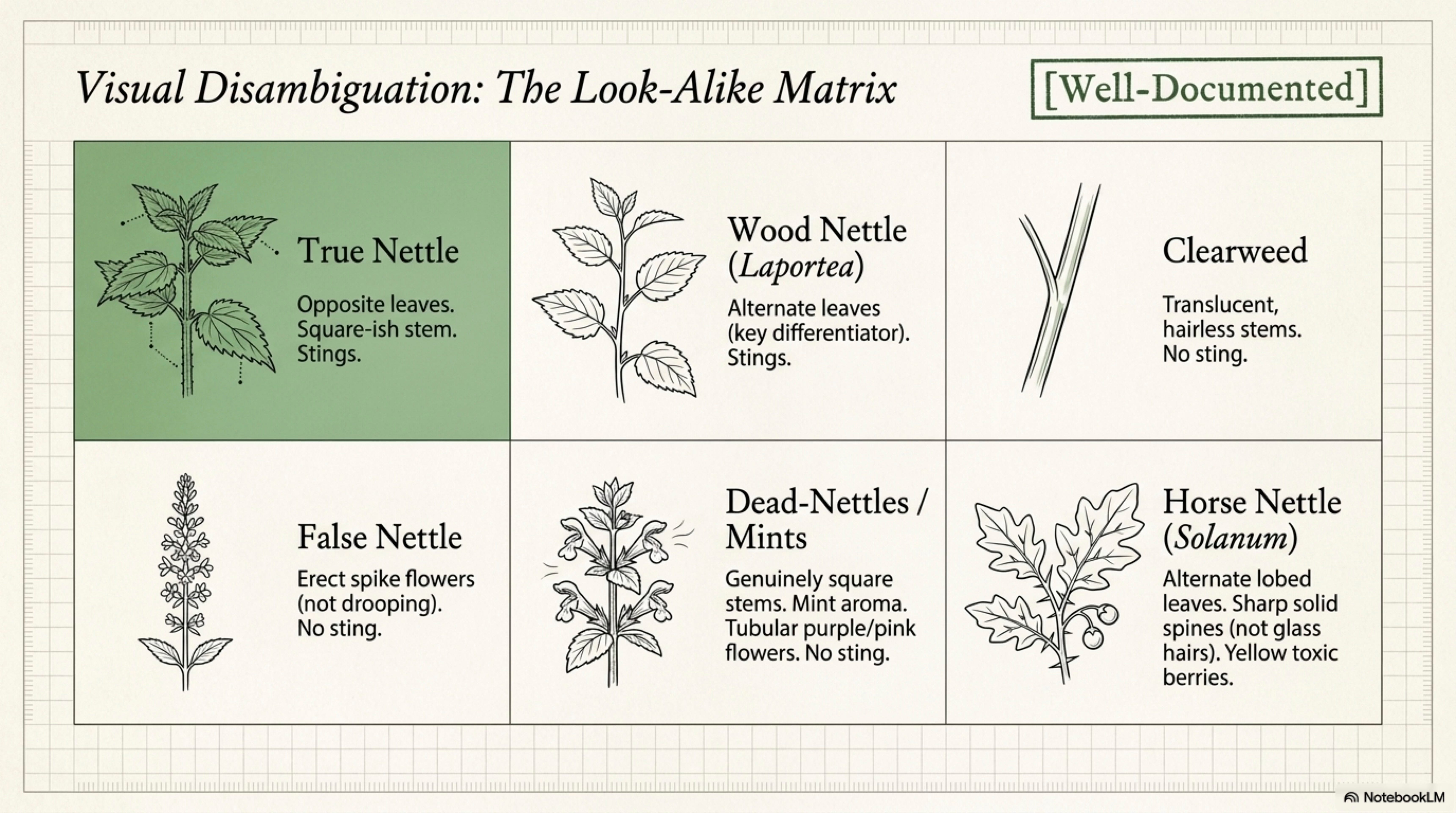
Wood nettle, Laportea canadensis (eastern N. America). Stings. Leaves are alternate, not opposite, the single clearest field mark. Stinging hairs longer and more dispersed. Grows in richer, shadier, moister forest than Urtica prefers [Boufford 1997]. [Well-documented]
Clearweed, Pilea pumila (eastern N. America). Does not sting. Translucent, almost watery stems and smooth leaves. Opposite leaves, but hairless. Often grows with wood nettle in damp shade [Boufford 1997].
False nettle, Boehmeria cylindrica. Does not sting. Opposite leaves but without trichomes; inflorescences in erect, spike-like clusters rather than the drooping axillary racemes of Urtica. Same family [Boufford 1997].
Dead-nettles, Lamium spp. (henbit, purple dead-nettle, white dead-nettle). Do not sting. These are mints (Lamiaceae): square stems (genuinely square, not “square-ish”), tubular zygomorphic flowers often pink/purple/white, opposite leaves. Aromatic when crushed, the mint cue is immediate. [Well-documented]
Horse nettle, Solanum carolinense. Different family (Solanaceae), unrelated. Alternate leaves, lobed, with sharp spines (not trichomes) on stems and leaf veins. Flowers star-shaped, purple-white. Fruits yellow berries. Toxic. The name is misleading and has caused misidentifications; the visual signature is unmistakable once known. [Well-documented]
Hemp and mulberry seedlings. Neither stings; neither has opposite serrated leaves on a clearly four-angled stem. The confusion is rare but occasionally reported.
3.3 Safety note
The misidentification risks with nettle are low in either direction: the sting confirms identity, and no dangerous herb resembles it closely enough to be accidentally consumed in its place. The real safety considerations are about handling the correctly identified plant, not about confusing it with something else. Those belong in Section 13 (Phase II).
One caveat. In New Zealand, the native tree nettle Urtica ferox produces a severely more potent sting than U. dioica, with documented human fatality [Connor 1977]. Travelers who “know nettles” from the Northern Hemisphere should treat U. ferox with much greater caution; the sister-species lesson does not transfer. [Well-documented]
4. Botanical Character and Life Cycle
Life-form. Herbaceous perennial. Aerial shoots die back each autumn; rhizomes overwinter and re-emerge. In long-settled patches, the rhizomatous clone can be decades old even when no single shoot is older than a year [Taylor 2009]. [Well-documented]
Architecture. Orthotropic (upright) aerial shoots rise from a plagiotropic (horizontally spreading) rhizome system at 5–15 cm depth. The rhizome is the persistent skeleton of the plant’s presence in a place; the shoots are seasonal expressions. A single rhizome fragment of a few centimeters, bearing a node, can regenerate a new clonal patch given moisture and nutrient supply, the plant exploits any disturbance that breaks up the rhizome mass [Taylor 2009]. [Well-documented]
Root strategy. Shallow and wide. Dense fibrous roots branch from the rhizome network in the top 20–30 cm of soil, concentrating where organic matter is richest. Nettle is not a deep-rooted plant. Claims of mineral mining from subsoil layers are not supported by root architecture or by any primary study I could locate [Taylor 2009; the “dynamic accumulator” claim traces only to grey-literature sources, Hamaker 1982; Kourik 1986]. [Anecdotal for the dynamic-accumulator framing; Well-documented for high foliar nutrient content on fertile sites.]
Clonal spread. Horizontal rhizome extension at rates of tens of centimeters to over a meter per growing season on productive sites; lateral edge advance is often most rapid into freshly disturbed or enriched ground [Taylor 2009]. In mature stands, clonal reproduction dominates over seed reproduction; in colonizing populations, seed is more significant.
Germination cues. Seeds require light for germination, a shallow burial stays dormant; disturbance that brings seed to the surface triggers the flush [Taylor 2009]. Cold stratification enhances but is not strictly required. Temperature optimum for germination is moderate, in the 15–25 °C range. [Well-documented]
Seed bank. Persistent. Seeds remain viable in buried soil for at least several years; Taylor (2009) cites studies reporting viability beyond five years in some soil conditions. The persistence is part of why nettle returns so reliably to disturbed sites even when no surface plants were visible for years.
Flowering sequence. In Britain, shoot emergence late February through April depending on latitude and season; vegetative dominance April–June; flowering June–August; seed set July–September; aerial senescence October–November [Taylor 2009]. In North America (U. gracilis), the equivalent arc runs roughly three to four weeks earlier in the Pacific Northwest lowlands, parallel to the Britain timing in the Great Lakes and Mid-Atlantic, and two to four weeks later at higher elevations and northern latitudes [USA-NPN records]. [Well-documented for Britain; Emerging for fine-grained North American phenology.]
Pollination. Wind-pollinated (anemophilous). The explosive stamen dehiscence mechanism, the stamens are held under tension in the bud and snap outward on maturation, releasing pollen in a visible cloud, is a small spectacle on warm still days in full flower. Female flowers are receptive to airborne pollen from neighboring plants; in gracilis monoecious populations, geitonogamy (self-pollination within a plant) is possible and likely occurs at nonzero rate [Taylor 2009; Bassett et al. 1974]. [Well-documented]
Seed dispersal. Mostly gravity and short-distance dispersal. Seeds do not have specific adaptations for long-distance dispersal; some evidence of endozoochory (seeds passing through animals) and epizoochory (sticking to fur); significant transport by water in riparian settings [Taylor 2009]. Human-mediated dispersal via agricultural traffic, contaminated seed, and soil movement is substantial where native and introduced populations co-occur. [Traditionally supported; detailed dispersal-distance studies are thin.]
Disturbance response. Strongly positive. Soil disturbance that fragments rhizomes and exposes seed both favor nettle unless the disturbance is severe enough to remove the soil seed-bank (deep scrape, pavement, deposition). The plant is a textbook competitor-ruderal, expressing more of each strategy by turns as conditions shift [Grime et al. 2007; Taylor 2009]. [Well-documented]
Successional role. Mid-successional. Nettle colonizes abandoned pasture, disturbed river terrace, and middens; it dominates for years to decades on fertile sites; it is eventually overtopped by shrubs and trees in closed-canopy succession unless recurring disturbance resets the stage [Rodwell 1991–2000; Taylor 2009]. In traditionally managed hedgerows and farmyard edges, where low-level disturbance is continuous, nettle can hold its dominance indefinitely. [Well-documented]
Longevity. Individual aerial shoots: one growing season. Individual ramets (root + rhizome + shoot system): several years to a decade. Clonal genet: theoretically unlimited where conditions persist; documented clonal patches in Britain exceed several decades [Taylor 2009]. [Traditionally supported for genet longevity, ramet turnover makes direct measurement hard.]
The rhizome’s memory. What this life-cycle pattern means, in a working landscape, is that a nettle patch tells you where the nitrogen has been pooling for a long time. The rhizome did not arrive yesterday. The aerial shoots are a signal the plant broadcasts each spring; the signal is readable because the underground network has been keeping records longer than the reader has been watching.
5. Ecological Intelligence
5.1 Soil relationships
Preferred conditions. Moist, well-drained, deep soils rich in available nitrogen and phosphorus. Slightly acid to calcareous; tolerates pH ~5.0–8.0 with optimum near neutral [Taylor 2009; Ellenberg 1988]. [Well-documented]
pH indicator. Ellenberg reaction value R = 7, base-rich to neutral, mildly calcareous leaning. Not strongly diagnostic on its own; nettle tolerates a broad pH range if fertility is adequate [Ellenberg 1988]. [Well-documented]
Mycorrhizal status. Facultatively non-mycorrhizal. Most surveys of U. dioica root systems have found no or very weak arbuscular mycorrhizal colonization [Harley & Harley 1987; Wang & Qiu 2006]. This is a genuine ecological finding, not a sampling artifact, and it is part of why nettle thrives on highly disturbed, highly fertile soils where mycorrhizal networks have been broken or where excess nutrients suppress the fungal partnership. There is one intriguing strand of evidence suggesting that the root-localized UDA lectin may itself inhibit mycorrhizal colonization in Urtica [cited in the phytochemistry literature via Peumans et al. 1984 follow-ups]. [Well-documented for non-mycorrhizal behavior; Emerging for UDA-inhibition hypothesis.]
Post-split, U. gracilis populations in North America have not been systematically surveyed for mycorrhizal status [Gap flagged].
Bacterial associations. No specific N-fixing symbiosis has been reported in Urtica dioica. The plant’s nitrogen economy runs on uptake, not fixation, which is why it requires already-enriched soil to thrive [Gap flagged for detailed rhizosphere microbiome studies].
Root exudate effects. Nettle rhizomes and roots release organic acids and other exudates that likely contribute to the rhizosphere’s distinct nutrient-cycling dynamics, but the chemistry of nettle rhizosphere exudation has not been characterized in the way that, for example, Secale cereale rhizosphere exudation has been [Gap flagged].
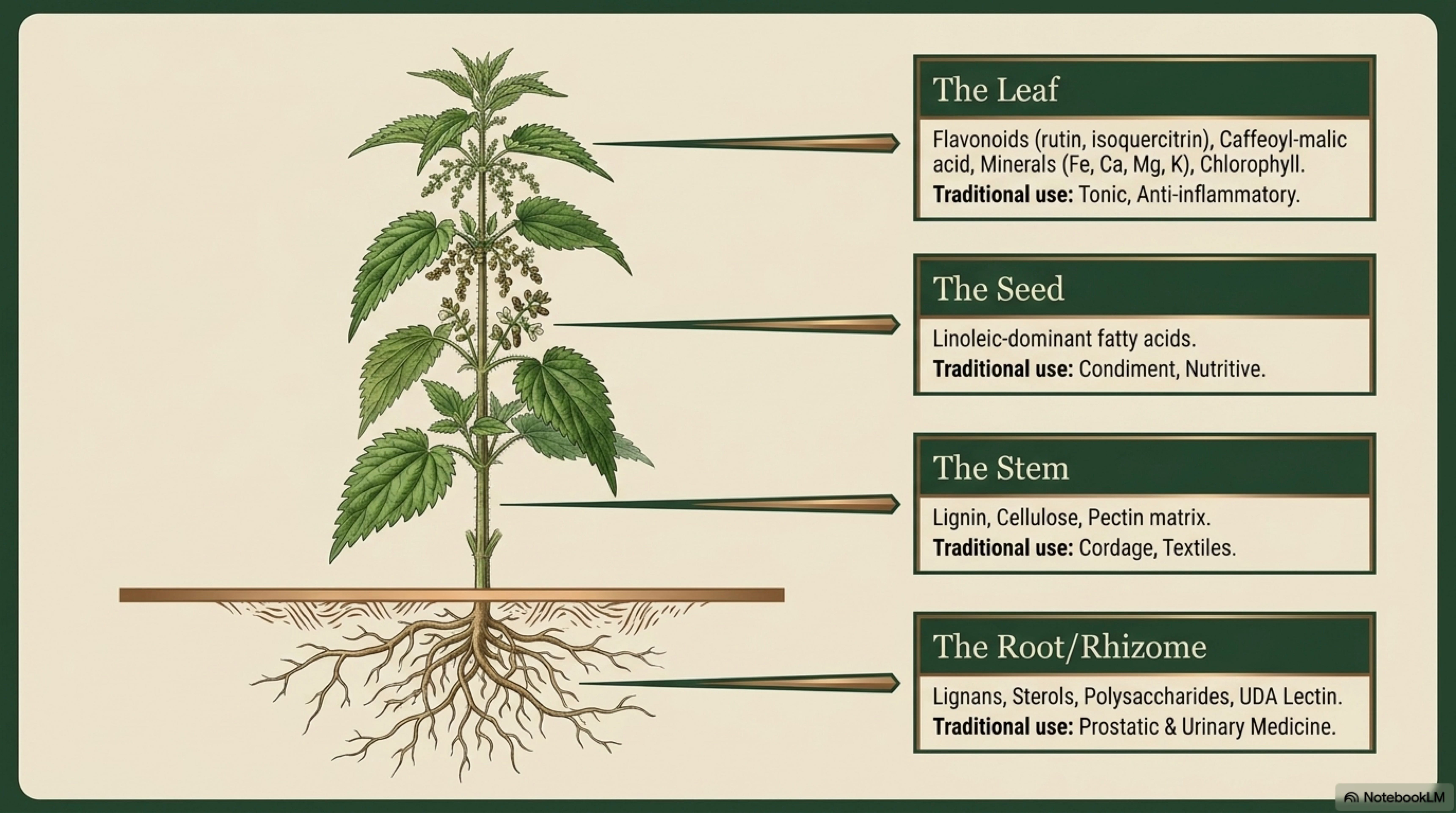
Nutrient accumulation. Foliar concentrations of N, P, K, Ca, Mg, Fe, and S in U. dioica are high compared to many temperate herbs [Taylor 2009; see Section 12 in Phase II]. This reflects high demand and high uptake from fertile substrate, not preferential extraction from deep or impoverished soil. The widely-circulated permaculture claim that nettle “dynamically accumulates” minerals by pulling them from depths other plants cannot reach is not supported by any primary study [tracing to Hamaker 1982 and Kourik 1986, neither of which presents experimental evidence]. Foliar nutrient analysis is real; the deep-mining narrative is an overreach. [Anecdotal for dynamic-accumulator framing; Well-documented for foliar content.]
Rhizosphere function. Dense root mats condition the upper soil horizons: aggregation, organic matter turnover, and macrofaunal habitat (earthworms in particular thrive in nettle-dominated soils, correlated with both the high nitrogen turnover and the disturbed-mesic conditions nettle favors). The specific microbial-community signature of nettle-dominated rhizospheres has not been characterized in a standardized way [Gap flagged].
Compaction implications. Nettle rhizomes can penetrate and fracture moderately compacted soils, and dense clonal patches tend to improve upper-profile friability over time. On severely compacted sites (pan layers, traffic zones), nettle tends to stay at the edges.
Allelopathy. No significant allelopathic effect on neighbors has been documented in Urtica dioica. The plant’s competitive dominance on fertile sites is better explained by fast growth, shade production, and high nutrient capture than by allelochemistry. The absence of allelopathy is itself noteworthy, many weedy plants of disturbed ground are allelopathic; nettle competes by outgrowing, not by poisoning. [Traditionally supported, absence of documented allelopathic literature despite significant community-ecology study.]
5.2 Water relationships
Moisture preference. Mesic to moist soils, Ellenberg F = 6. Thrives in a wide moisture band but struggles at extremes [Ellenberg 1988; Taylor 2009]. [Well-documented]
Drought. Aerial shoots are drought-sensitive, moderate summer drought wilts nettle visibly within days and can kill shoots back to the rhizome. Rhizomes themselves are surprisingly drought-tolerant and re-sprout readily when moisture returns [Taylor 2009]. The plant reads drought as a signal to retreat, not die.
Flood tolerance. High. Nettle tolerates weeks of partial submergence on floodplains during dormancy and during early growing season; rhizomes handle anoxia better than shoots [Taylor 2009]. Riparian nettle stands on seasonal floodplains are one of the most reliable nettle habitats in temperate Europe and North America.
Water-table association. Fertile seasonally wet meadows, wet woodland edges, riverside hedgerows, and floodplain terraces are classic nettle habitat [Rodwell 1991–2000]. Nettle does not tolerate permanent saturation, it is not a true wetland plant, but seasonal high water tables with summer draw-down suit it well.
Riparian role. Dense rhizome mats likely contribute to bank stabilization on floodplain edges and shallow river terraces. The quantitative evidence for this (erosion-pin studies, bank-shear measurements) is thin; the claim is widely repeated and plausible rather than formally demonstrated [Gap flagged]. [Traditionally supported]
5.3 Community ecology
Companion plants. Classic nettle-dominated communities in Britain include Galio-Urticetea (Urtica-Galium cleaver associations), NVC OV24 (urtico-galietum aparines), and nettle-rich phases of W8 and W10 woodland communities [Rodwell 1991–2000]. In working farmland, nettle pairs reliably with cleavers (Galium aparine), ground elder (Aegopodium podagraria), herb bennet (Geum urbanum), and hedge garlic (Alliaria petiolata), a suite of nitrogen-enrichment specialists [Taylor 2009]. [Well-documented]
Competitive behavior. Strongly competitive on its preferred sites. Once established on fertile moist ground, nettle produces dense shade, high biomass, deep litter, and a self-reinforcing nitrogen-rich microenvironment that excludes slower-growing herbs. Grime’s CSR classification places U. dioica firmly in C-strategist territory, tall, leafy, fast-growing, with high resource demand [Grime et al. 2007]. [Well-documented]
Nurse functions. Nettle does not function as a nurse plant in the classical sense (providing shelter for slower establishment of woody pioneers); it tends rather to delay succession by dominating the herb layer for decades.
Wildlife value. Very high. Four specialist butterfly species (see 5.4 below) depend on nettle as larval host in Britain; the same pattern holds in continental Europe with some substitutions; North American native nymphalids similarly use U. gracilis [Dennis 1992; Scott 1986]. Nettle stands support spider assemblages, ground-beetle communities, and small-bird foraging (wrens, warblers) at notably high densities [Taylor 2009]. [Well-documented]
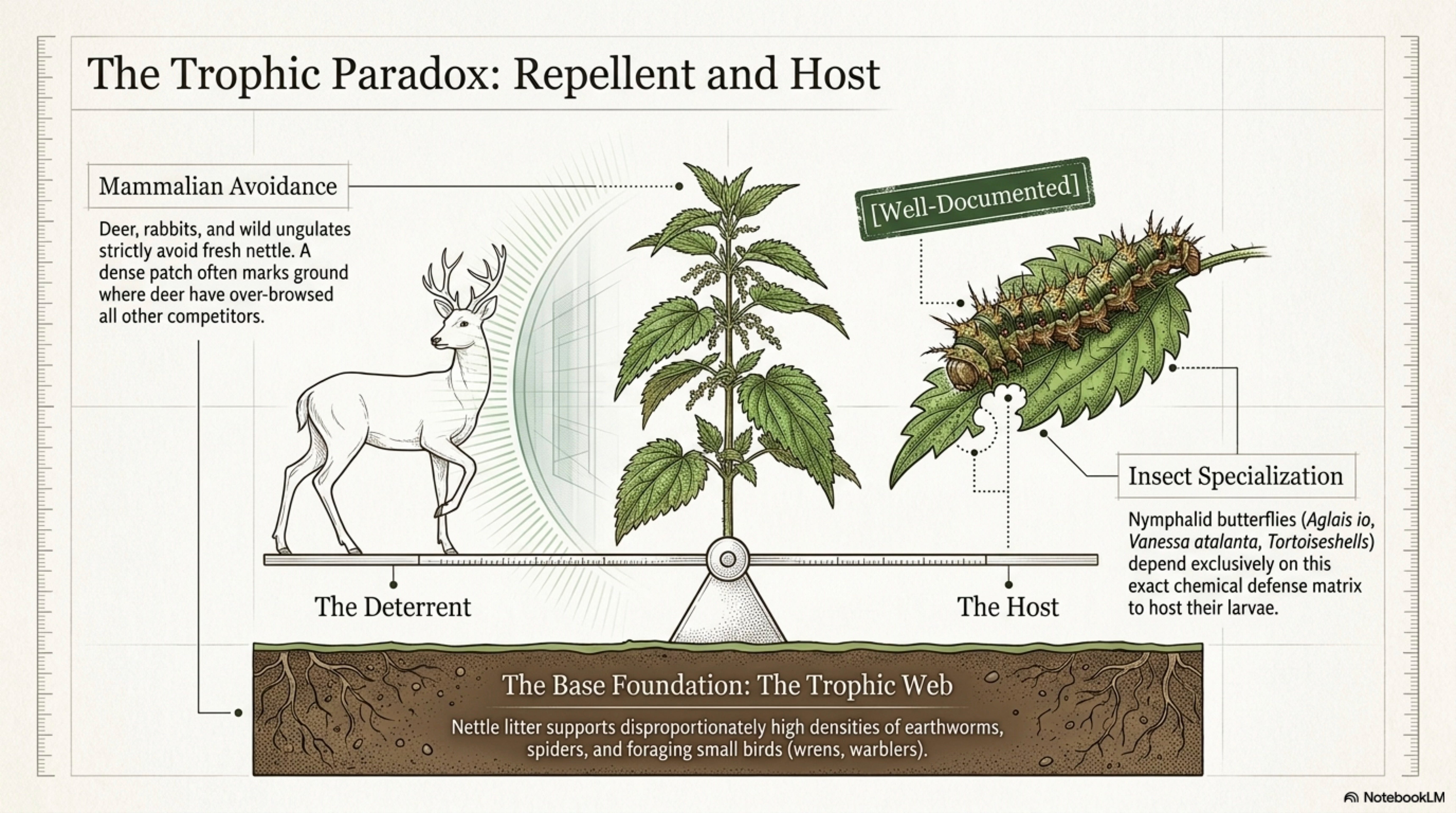
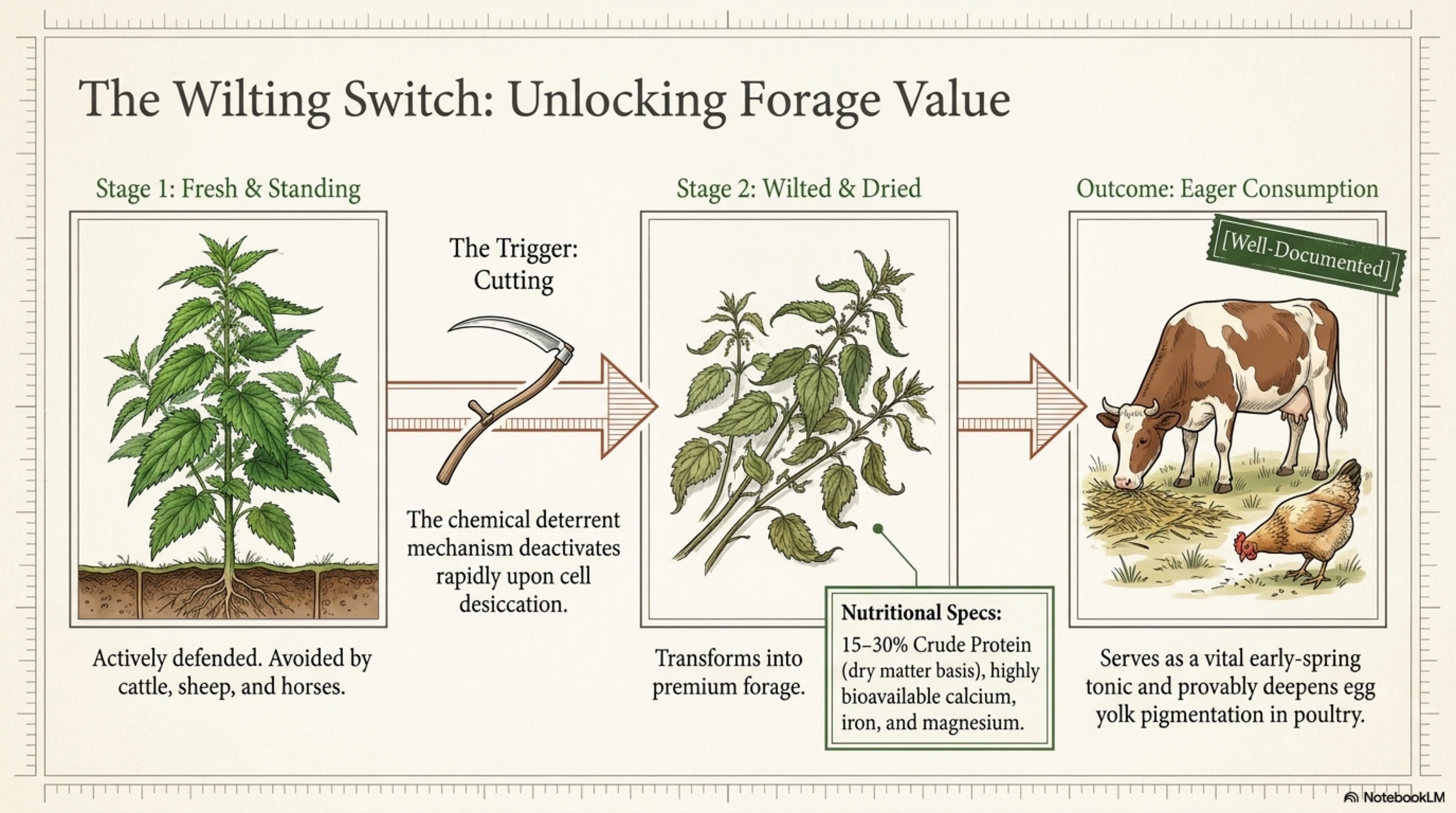
Herbivore relationships. Deer, rabbits, sheep, and cattle all avoid fresh nettle. The sting is a deterrent that works. Wilted or dried nettle is readily eaten by sheep, cattle, pigs, and poultry [see Section 6]. The deterrent/palatability switch on wilting is one of the most exploitable facts about the plant. [Well-documented]
Habitat role. In riparian woodland, farmyard edge, hedgerow, and recovering waste ground, nettle stands function as long-duration nitrogen reservoirs, invertebrate habitat, and structural refuge. In undisturbed old-growth forest and in nutrient-poor acidic moorland, nettle is absent, it is a plant of disturbance and enrichment, not a generalist.
5.4 Pollinators and insects
Pollinator value. Wind-pollinated; flowers offer no nectar and are not insect-attractive. Bees and other pollinators do not visit nettle for floral resources [Taylor 2009]. The plant’s insect-ecology contribution runs almost entirely through larval host relationships, not through pollination services.
Larval host relationships, a textbook specialization.
In Britain and much of Europe, four butterflies of the family Nymphalidae use U. dioica as larval host: Aglais io (peacock), Aglais urticae (small tortoiseshell), Vanessa atalanta (red admiral), and Polygonia c-album (comma) [Dennis 1992]. The tortoiseshell’s specific epithet urticae is itself a naming of the relationship. These species’ adult dispersal patterns are constrained by the distribution of nettle patches large enough and nitrogen-rich enough to support larval development [Pollard 1979]. [Well-documented]
Nettle chemistry, the same trichome cocktail, oxalate crystals, and flavonoid-tannin complex that deters mammalian herbivores, has apparently been overcome evolutionarily by these four species, which sequester or tolerate the defensive compounds and in some cases use them for their own defense. Aglais urticae larvae preferentially oviposit on regrowth from clipped or mown nettle, evidently responding to higher tissue nitrogen and lower chemical defense in young regrowth [Pullin 1987]. This is an applied fact: managed, cut-and-regrow nettle patches produce more butterflies than unmanaged old stands. [Well-documented]
In North America, the analogous specialists include Vanessa atalanta (circumpolar), Polygonia satyrus, Aglais milberti (Milbert’s tortoiseshell), and various Nymphalis spp., all using native U. gracilis [Scott 1986]. The pattern of nymphalid–Urtica specialization is a cross-continental phenomenon, not an artifact of Eurasian biogeography. [Well-documented]
Insectary value. Nettle patches host predatory spider, beetle, and wasp communities at high densities [Taylor 2009]. The aphid Microlophium carnosum feeds on nettle and in turn supports ladybird, lacewing, and hoverfly larvae, nettle stands near orchards and gardens function as beneficial-insect reservoirs [British Trust for Ornithology and allied extension guides]. [Well-documented]
Nectar and pollen timing. Not relevant for pollinators, but the wind-pollen cloud of nettle in June is a documented hay-fever contributor in sensitive individuals [Taylor 2009].
Overwintering relevance. Standing dry nettle stems overwinter invertebrate communities, including overwintering stages of the nymphalid specialists [Dennis 1992]. Late-autumn clearance of nettle patches on working farms is a documented negative impact on butterfly populations.
Beneficial predator support. High. Nettle is a textbook “beneficial insect refuge” in IPM literature, often recommended in orchard and field-margin plantings [extension literature; e.g., Noble Research Institute and European equivalents]. [Well-documented]
5.5 Ecosystem functions
Soil building. High-N litter with moderate C:N (~15–25) decomposes quickly. Nettle stands cycle nitrogen and phosphorus aggressively, increasing topsoil organic matter and contributing to the characteristic dark, friable, earthworm-rich surface of long-established patches [Taylor 2009; Grime et al. 2007]. [Well-documented]
Carbon contribution. Modest to moderate per unit area. Nettle’s biomass turns over rapidly, fast-decay litter does not build long-term carbon stocks the way slower-decay grasses or woody plants do. Net carbon contribution per hectare is real but not exceptional.
Erosion control. Dense rhizome mats likely provide significant topsoil stabilization on disturbed ground. Quantitative studies specific to Urtica are thin [Gap flagged]. On riparian banks, nettle contributes to a broader cohort of mesic-soil-stabilizers (cleavers, ground elder, rough meadow-grass, etc.).
Shade and shelter. Stands 1–2 m tall produce dense shade by mid-season, creating cool moist microclimates exploited by amphibians, small mammals, and ground-dwelling birds.
Biodiversity support. Disproportionately high in Nymphalidae, spiders, and soil fauna, especially earthworms [Taylor 2009; Dennis 1992].
Restoration. Useful as an interim cover on nitrogen-loaded disturbed ground, recovered brownfield sites, grazed-out pasture corners, post-flood riparian terraces. Native-species restoration projects frequently regard nettle as an expected intermediate phase to be worked with rather than eradicated.
Phytoremediation. U. dioica shows moderate tolerance and accumulation of Cd, Zn, Pb, and Cu on contaminated soils [Grejtovský et al. 2006]. It is not a hyperaccumulator; it functions reasonably as a bioindicator of heavy-metal contamination and as a phytostabilizer on moderately contaminated sites. Practical remediation roles are limited to moderate-contamination conditions. [Well-documented for tolerance and moderate accumulation; Emerging for practical application.]
5.6 Indicator value
Nettle is one of temperate Europe’s most diagnostic indicator plants. Each line below is a specific signal the plant sends.
Fertility. High. Nettle dominance indicates soils rich in available N and P; it is a Top-5 indicator of nitrogen-enriched conditions in Ellenberg systems [Ellenberg 1988]. [Well-documented]
Compaction. Variable. Nettle can establish on moderately compacted ground but abandons severely compacted sites; clonal expansion tracks friable, biologically active profiles.
Disturbance history. Strong positive indicator of disturbance within the last decades. Old-growth forest understory is not nettle habitat; farmyard, pasture edge, and riparian terrace is.
Successional stage. Mid-successional; ruderal-competitor. Indicates the ecosystem is past bare-ground colonization but has not yet reached closed woody canopy.
Moisture. Mesic to moist; avoids true wetland and true drought.
Salinity. Intolerant, absent from salt marsh and saline prairie.
Contamination. Tolerates and mildly accumulates several heavy metals; patchy presence on mine-spoil and contaminated industrial ground [Grejtovský et al. 2006]. [Well-documented]
Grazing pressure. Indicates overgrazed or dung-patch-enriched pasture on productive soils; cattle and sheep grazing around middens and camp corners increase nettle abundance.
Microbial imbalance. Not a specific indicator. Nettle’s non-mycorrhizal habit means its dominance can signal sites where mycorrhizal networks have been disrupted, but the correlation is weak [Emerging].
Mineral deficiency or excess. High foliar content reflects substrate fertility, not substrate imbalance. Nettle is not a reliable diagnostic for trace-element deficiency or toxicity.
5.7 Ecological synthesis
Watch where the nettle grows. Not the scattered seedlings of disturbance, the dense stand, shoulder-high by midsummer, crowding the fence line below the cow-camp and the hedge base where the dog-fox beds each May. The stand tells you something the soil would otherwise keep to itself: that for years, maybe decades, nitrogen and phosphorus have pooled here. That disturbance has recurred often enough to keep trees from closing in. That earthworms have worked the top six inches into a dark friable tilth that retains moisture through August. That the moisture comes seasonally and leaves seasonally, so the plant can rest its rhizomes through a wet spring and lift its shoots through a dry summer. That no salt has reached here and no pan of compaction has set deeper than plow depth. That the soil remembers livestock and human presence, and that the remembering has been fed forward, year by year, by the plant itself. Nettle is not a sign of neglect, nor of abundance alone. It is a sign of a place where humans and animals have been, and where the ground has been fed more than it has been stripped. The rhizome keeps the ledger; the shoots announce the accounts. When you see a mature nettle patch holding its line against cleavers and ground elder, what you are seeing is the slow geological work of a single chemistry, a plant that metabolizes settlement itself into biomass, pigment, fiber, and medicine, and hands the ledger back to the soil with interest.
6. Animal Interactions and Ethology
Animals are teachers. Nettle’s relationships with non-human life are an axis of knowing as old as the plant, older than any human materia medica.
6.1 Wild animal relationships
Mammals that browse. Very few fresh-nettle browsers. Deer (roe, red, white-tailed, mule), rabbits, hares, and most wild ungulates avoid fresh U. dioica except in severe winter shortage [Taylor 2009]. The sting works as a mammalian deterrent; it does not dissuade specialist insects. [Well-documented]
Mammals that avoid. The primary pattern. In long-term exclosure studies on European woodland, nettle dominance tracks inversely with mammalian browsing pressure on competitor species; heavy deer herbivory on preferred herbs allows nettle to expand [Taylor 2009]. A nettle stand can mark a place where the deer have eaten everything else.
Bird relationships. Wrens, warblers (garden warbler, blackcap), and other small insectivores forage intensively in nettle stands for caterpillars and aphids [British Trust for Ornithology observations]. Robins and thrushes occasionally take seeds. Pheasants and pigeons take nettle seeds in autumn. No bird is a specialist on nettle, but several rely on nettle-hosted invertebrates. [Well-documented]
Reptile/amphibian. Nettle stands offer cool moist shelter for slow-worms, common lizards, and frogs; on riparian terraces, amphibian densities can be high under dense nettle cover [Taylor 2009]. [Traditionally supported]
Insect beyond pollination. See 5.4 above. The nymphalid specialization is the flagship story. Beyond butterflies: the nettle aphid Microlophium carnosum, the nettle weevil Phyllobius pomaceus, and several hemipteran and dipteran associates [Taylor 2009]. [Well-documented]
Soil fauna. Earthworms at notably high densities in nettle-dominated soils [Taylor 2009]; Collembola, mites, and isopods at high-fertility levels under nettle litter. The plant’s high-N fast-decay litter supports decomposer communities intensively. [Well-documented]
6.2 Zoopharmacognosy
Documented self-medication, thin and contested. No peer-reviewed zoopharmacognosy study of specific nettle-seeking behavior in wild or domestic animals has reached the strength of, for example, the chimpanzee Aspilia literature [e.g., Huffman 1997]. Horse-owners and goat-graziers commonly report that animals will seek out and eat nettle, wilted or standing, at particular times of year, often in early spring after winter confinement; the observation is widely repeated but has not been formally studied. [Anecdotal]
Correlation with known pharmacology. If the anecdotal reports are accurate, the correlation to the nutritional literature (very high protein, iron, calcium, magnesium, see Section 12) and to the anti-inflammatory/anti-allergic literature would be consistent with an animal-mediated recognition of early-spring tonic value. [Frontier Hypothesis, see §20.]
Veterinary ethnobotany. Traditional European livestock practice has long included dried nettle as a winter/spring tonic for horses, cattle, pigs, and poultry; the evidence base is agronomic rather than zoopharmacognosy-experimental. [See Section 6.3 and Phase II §14.] [Traditionally supported]
6.3 Livestock relationships
Forage value (summary).
Crude protein: very high. Aerial parts of U. dioica routinely report 15–30% CP on a dry-matter basis across growth stages; young pre-flowering shoots at the upper end of this range
Fiber (NDF/ADF): moderate.
Minerals: iron, calcium, magnesium, potassium notably high.
Chlorophyll / carotenoids: high; yolk-pigmentation effect in poultry is well-documented.
Anti-nutrients: oxalate; cystolith formation post-flowering affects palatability and mineral bioavailability. [Well-documented]
Palatability by species.
Cattle: avoid fresh; readily eat wilted or ensiled.
Sheep: avoid fresh; readily eat wilted, dried, or as hay mixed component.
Goats: some browse fresh young shoots; readily eat wilted.
Horses: generally avoid fresh unless severe shortage; readily eat dried as tonic.
Pigs: eagerly eat fresh cut-and-wilted; historical pig-feeding literature is extensive.
Poultry (chickens, ducks, geese): readily eat chopped fresh, wilted, or dried. Effects on yolk color and egg quality documented. [Taylor 2009; Kara 2016; see Phase II §14 for numbers.]
Behavioral indicators. Livestock avoidance of fresh nettle is a reliable pasture signal: where nettle dominates, grazing pressure has been either absent or so severe on surrounding forage that nettle filled the gap. Conversely, concentration of nettle around gate-corners, water-troughs, and night-yard areas is a dung-enrichment marker, the pattern is so reliable that it can diagnose livestock movement in old fields.
Milk/egg/meat quality. Dried nettle inclusion in dairy cattle and laying-hen rations is documented to affect butter color (greener-yellow), yolk pigmentation (deeper orange), and in some studies lay-rate and feather quality [Loetscher 2013; see Phase II §14]. [Well-documented]
6.4 Animal-plant-soil feedback loops
Grazing effects on plant chemistry. Clipping and regrowth alter leaf chemistry: regrowth tissue is lower in fiber, higher in soluble protein, and sometimes lower in defensive compounds, precisely why Aglais urticae prefers regrowth for oviposition [Pullin 1987]. [Well-documented]
Seed and nutrient distribution via animals. Endozoochory of nettle seeds is limited; epizoochory (on fur) occurs but is not a major dispersal mode. The more important animal-mediated effect is nitrogen concentration: dung, urine, and carrion enrich patches around which nettle then establishes and expands. [Traditionally supported; dispersal-distance studies thin.]
Dung/urine interactions. Nettle responds positively to recent dung deposition with rapid growth and often visibly brighter-green foliage. Urine patches with very high local N can kill fresh nettle, then become nettle-dominated on recovery as surrounding soil re-equilibrates.
Managed grazing implications. In regenerative grazing systems, dense nettle patches signal either rest-phase (fertility built up without recent disturbance) or a camp/corner accumulation zone. Mob-grazing with high stock density followed by long rest can reduce nettle dominance on productive pasture by restoring herbage competition and by mechanical trampling that weakens rhizomes. Under-grazing or continuous low-density grazing tends to let nettle expand.
6.5 Animal interaction synthesis
The deer do not touch the plant. The cow refuses it standing, then eats it wilted at sundown. The small tortoiseshell lays her eggs on the regrowth of a sheep-clipped patch, twice as many larvae on the cut strip as on the uncut border three meters away, because the tissue chemistry shifted and the butterfly read the shift. The wren dives into the stems at the top of the hedge, takes a larva, feeds a chick. The earthworms move up toward the surface under nettle litter because the C:N is right and the moisture holds. A single plant binds a mammal that avoids it, an insect that depends on it, a bird that feeds from it, a decomposer that thrives beneath it. What the animals know, long before any herbal is written, is that this plant is a concentrator, of nitrogen, of protein, of pigment, of a particular kind of fast-decay energy. The sting is the plant’s way of choosing its partners. Those who cannot handle it pass by. Those who can, the nymphalid larva with its tolerant gut, the cow with its wilting patience, the horse that learns to bite past the tip, the human who learns to wear sleeves and harvest in the morning when the trichomes are fullest and the cuticle most brittle, these are the kin the plant has courted across millennia, and the network of relationships is itself the sign of what the plant is actually for.
7. Climate Resilience and Adaptation
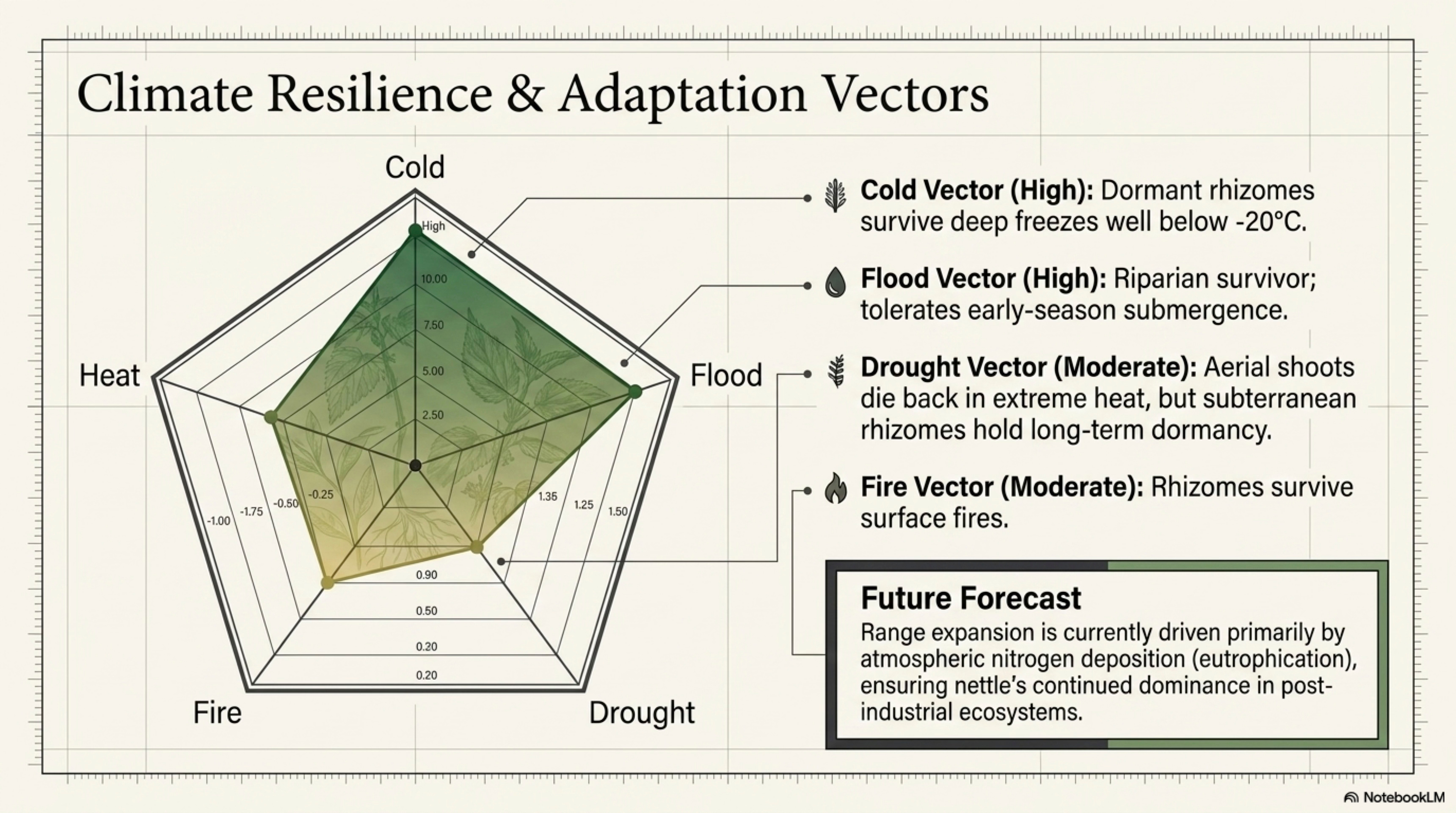
Heat tolerance. Moderate. U. dioica tolerates short summer heat spells when soil moisture is adequate; prolonged heat combined with drought causes shoot desiccation and rhizome dormancy until moisture returns [Taylor 2009]. The plant is a temperate-maritime species; true continental summer extremes limit its distribution. [Well-documented]
Cold tolerance. Very high. Rhizomes survive soil freezing to well below −20 °C in dormancy [Taylor 2009]. Northern range limits in Europe and North America are set more by summer temperature (season length for flowering and seed set) than by winter cold. [Well-documented]
Drought tolerance (soil moisture). Moderate at the rhizome level; poor at the shoot level. Wilt-point shoots re-sprout from rhizomes when moisture returns, so the clonal population survives droughts that kill individual shoot cohorts. [Well-documented]
Flood tolerance. High during dormancy and early growing season (see §5.2). Nettle is one of the reliable components of floodplain herb communities in Europe and North America. [Well-documented]
Fire tolerance. Low to moderate. Fresh green stands carry fire poorly (high moisture content); dry late-season stands and litter can carry surface fire; rhizomes generally survive surface fires and re-sprout in the following season. Nettle is not a fire-adapted plant but it is not eliminated by low-intensity surface fire. [Traditionally supported]
Salinity. Intolerant. Absent from salt-marsh, salt-steppe, and coastal brackish communities [Ellenberg 1988]. [Well-documented]
Wind tolerance. Moderate. Tall standing crops can lodge in heavy wind, particularly late in the season when stems become fibrous and top-heavy with seed. Lodged plants usually re-establish upright growth if still early enough in season.
Plasticity. Extensive. Nettle expresses phenotypic plasticity in height (30 cm to >2 m depending on nutrient and moisture regime), leaf size, trichome density, and flowering timing. This plasticity is part of why a single species concept spans such a range of habitats and why the taxonomic treatment of subspecies and segregate species has been historically contested [Taylor 2009].
Observed and projected range shifts. Nettle is expanding in much of Europe, driven primarily by atmospheric nitrogen deposition rather than by temperature [Pitcairn et al. 1998; Bobbink et al. 2010]. Range limits are shifting poleward at modest rates consistent with general climate-warming range shifts, but the dominant driver of nettle abundance trends is eutrophication, not warming per se. [Well-documented for Europe; Gap flagged for comparable analyses on North American U. gracilis.]
Future regenerative relevance. Because nettle thrives on nitrogen-enriched disturbed ground, it is likely to remain or expand in the ecosystem assemblages produced by agricultural intensification, climate-driven extreme-weather disturbance, and post-abandonment recovery of former pasture and cropland. For regenerative practitioners, this means nettle will increasingly be a plant to work with rather than a plant to try to eliminate, a free-of-charge protein crop, forage resource, fiber source, and ecological amenity on ground that other crops would require substantial amendment to support.
8. Phenology and Working Calendar
8.1 Seasonal cycle
Britain-centered timing; adjust for latitude and local microclimate. Pacific Northwest lowlands run ~3 weeks ahead; continental interior east of the Rockies runs roughly on UK timing; higher elevations and northern latitudes run 2–4 weeks behind.
Emergence: late February to April. Earliest shoots often appear in sheltered south-facing hedge-bases and warm riparian corners weeks before general emergence. [Taylor 2009; Woodland Trust Nature’s Calendar]
Vegetative dominance: April–June. Rapid height extension, leaf expansion, peak chlorophyll. This is the window for food, medicine, and fiber-precursor harvest.
Flowering: June–August, with variation by latitude and genotype. Anemophilous pollination; explosive stamen release most visible on warm still days in full flower.
Seed set: July–September. Pendulous female inflorescences become loaded with small brown achenes; nutrient translocation from leaves to seed accelerates.
Aerial senescence: October–November. Leaves yellow, drop; stems stand through winter in many sites, weathering and retting in place.
Dormancy: November–February. Rhizomes overwinter; buds set close to the soil surface. [Well-documented]
8.2 Timing triggers
Day length: less critical than for many herbaceous perennials; emergence is temperature-triggered more than photoperiod-triggered within the Holarctic range [Taylor 2009].
Temperature thresholds: emergence typically begins when mean soil temperature at 5 cm exceeds ~5 °C sustainedly; accelerating growth above 10 °C.
GDD (growing degree days): flowering typically requires accumulation of ~900–1200 GDD base 5 °C from emergence in temperate European populations, approximate figures with genotype and site variation [Traditionally supported; no standardized published GDD study for U. dioica specifically located].
Rainfall: spring moisture accelerates shoot extension; summer drought compresses the vegetative window.
Traditional seasonal markers: “nettle out, pigeon in” (rural English); first nettle harvest at the Celtic festival of Imbolc (early February) in mild years; Scandinavian nässelsoppa at the spring equinox; Greek Orthodox Lenten horta tradition timed to early-spring emergence. [Traditionally supported]
Companion plant cues: nettle emergence typically coincides with celandine (Ficaria verna) flowering, bluebell leaf-up, and early hedge blackthorn bud break in British populations.
8.3 Practical working windows
Leaf harvest for food and fresh medicine: from first 4–6 inches of shoot to just before flowering. The traditional European and Indigenous North American rule, don’t eat nettle after it flowers, is grounded in cystolith formation (calcium carbonate crystals that develop as leaves mature) and in mild gastrointestinal irritation reported from post-flowering leaves [Traditionally supported across European folk, Pacific NW Coast, and Chinese sources].
Seed harvest: late summer to early autumn, when pendulous female inflorescences are heavy and brown but before shatter.
Root harvest for medicine (BPH, diuretic): autumn or early spring when rhizomes are carbohydrate-rich, after aerial die-back or before full shoot expansion.
Fiber harvest: late summer to autumn, when stems are fully elongated and bast fiber is mature but before heavy winter weathering.
Compost-activator / biodynamic prep 504 harvest: full flowering stage (June–July in much of Europe).
Fermentation (purin d’ortie, FPJ, lacto-ferment): pre-flowering vegetative stage, when leaf chemistry is at peak nutritional density.
Propagation (rhizome division): dormant season (late autumn to early spring) or after first flush, with moisture.
8.4 Sensory timing notes
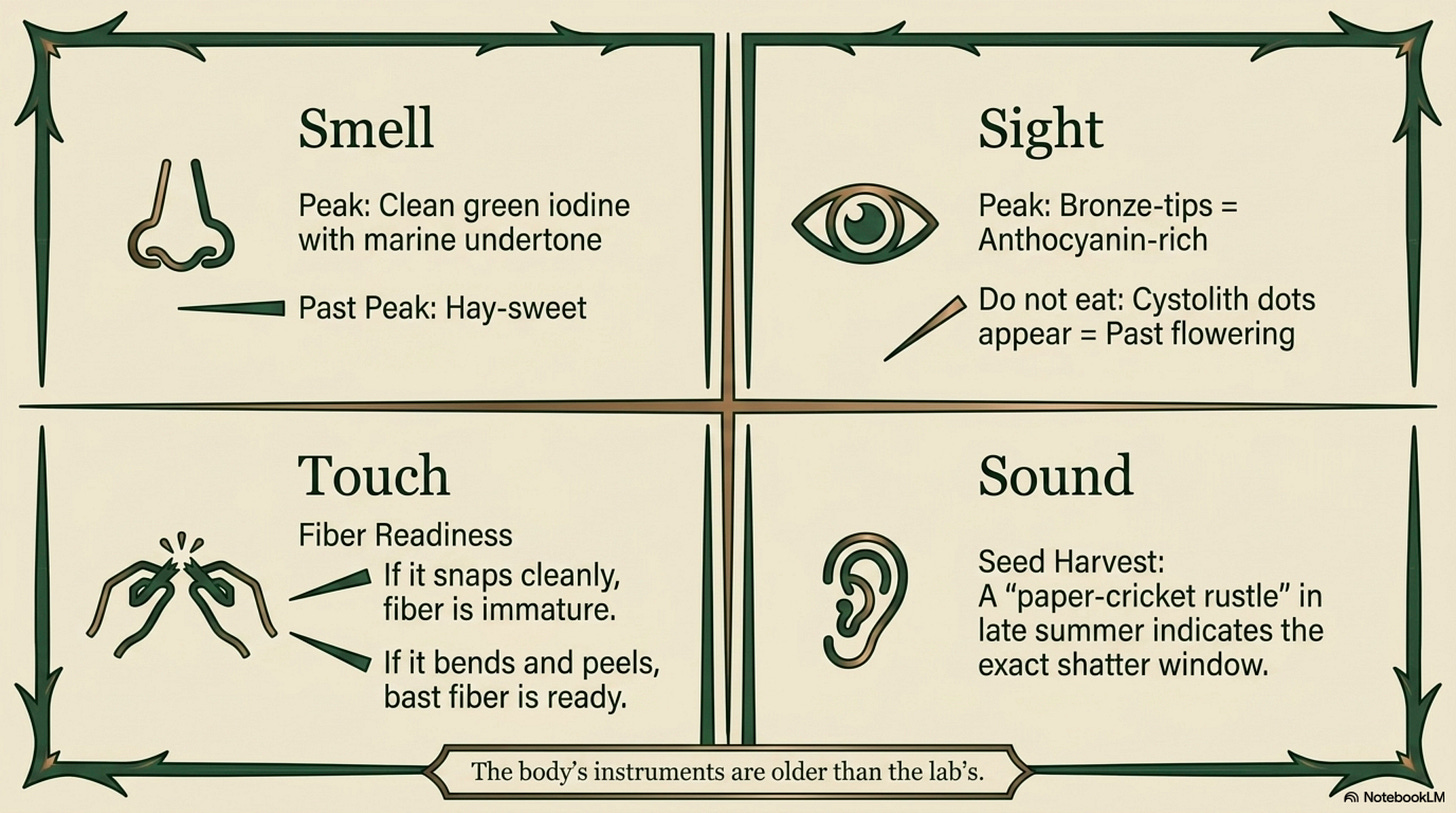
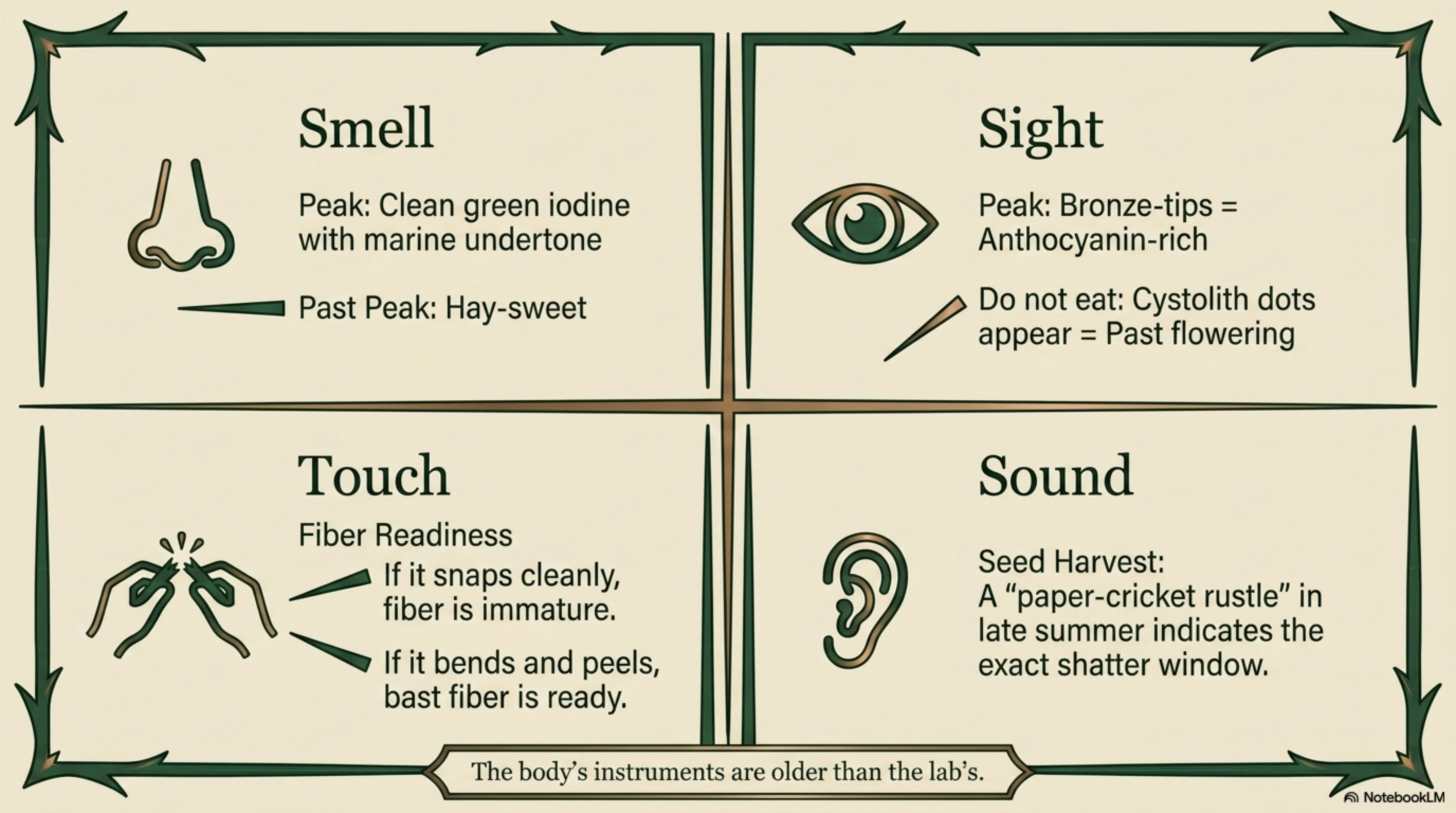
Field knowledge. The body’s instruments are older than the lab’s.
Aroma at peak: mid-morning on the first warm day of April, fresh young shoots crushed between finger and thumb, a clean green iodine-like note with a marine undertone. If the smell has turned strongly hay-like without warmth behind it, the peak harvest window has closed.
When bitterness changes: late June, when flowering begins, leaves shift from sweet-grassy to distinctly astringent. A single leaf tried raw (cautiously, or blanched) tells you whether the patch is still in culinary window.
When tissues become fibrous: stems past flowering can be snapped cleanly only at the lower nodes; the upper stem resists breaking and begins to peel. This is the transition signal for fiber harvest, if the stem bends before it breaks, the fiber is forming well.
Insect activity signals: if the patch is crowded with small tortoiseshell or peacock larvae, the plant is in active nymphalid production; clipping is a decision to weigh against the lives in the stand.
Color changes indicating chemistry: young leaves with a bronze tint at the tip carry higher anthocyanin, common in cold-stressed early-spring shoots and often considered strongest tonic material by folk practitioners.
When the plant “tells you” it’s ready: the patch has shifted from soft new green to full dark green; stems are upright and firm; leaves have reached full size but not yet begun to dull; pollen has not yet released. This is the narrow window, typically 2–3 weeks per patch per year, when food, fresh medicine, and fiber-precursor harvest all align. The rural European phrase for this window is simply “nettle time.”
9. History, Folklore, and Cultural Memory
9.1 Historical timeline
Bronze Age, textile signal. The Lusehøj burial textile (Voldtofte, Denmark, ~800 BCE) was long assumed to be flax. Bergfjord et al. (2012) identified it by polarized-light microscopy and calcium-oxalate signature as nettle, and, more striking, showed by strontium isotope analysis that the fiber was imported from the Kärnten-Steiermark region of the Austrian Alps, not local Danish nettle [Bergfjord et al. 2012]. Bronze Age nettle cloth was sufficiently valued to move across Europe. Neolithic Swiss lake-dwelling sites (e.g., Arbon Bleiche 3) have Urtica achenes in macrofossil assemblages, with dietary and fiber use ambiguous [Jacomet 2006]. [Well-documented]
Classical period. Pliny (NH XXII.13–17, 1st c. CE) records nettle as food, medicine, and urtication agent; Dioscorides (IV.93) codifies the pharmacognosy that European herbalism will repeat for sixteen centuries, hemostatic for nosebleed and wound, diuretic, emmenagogue, rheumatic. Virgil mentions nettles in fodder context. Galen classifies the plant as hot and dry in the second degree, diuretic and resolvent. [Well-documented]
Anglo-Saxon and medieval. The Lacnunga manuscript (Harley MS 585, 10th–11th c.) names wergulu, nettle, as sixth of the Nine Herbs against “flying venom” and infection [Pettit 2001]. Bald’s Leechbook (BL Royal 12 D xvii, 9th–10th c.) uses nettle in wound poultices and in drinks against “elf-disease” [Cockayne 1865]. Hildegard of Bingen (Physica I.87, c. 1150) prescribes spring nettle to purge phlegm from the stomach and warm cold constitutions [Throop 1998]. Strabo’s Hortulus (9th c.) and Macer Floridus’s De Viribus Herbarum carry the same Dioscoridean core forward. The Trotula texts of 12th-century Salerno incorporate nettle seed into gynecological formulations [Green 2001]. [Well-documented]
Early modern. Gerard’s Herball (1597), Parkinson’s Theatrum Botanicum (1640), and Culpeper’s Complete Herbal (1653) expand the medieval synthesis with regional English additions. Culpeper’s “Mars owns the herb” assigns nettle an astrological signature that downstream Western herbalism still invokes. Fuchs (1542) and Bock (1539) introduce German vernacular knowledge; Dodoens (1554) codifies the Dutch tradition. [Well-documented]
19th and early 20th century. Maud Grieve’s A Modern Herbal (1931) compiles Western European nettle knowledge into the single most cited modern reference. Samuel Thomson’s American Eclectic tradition treats nettle as alterative and diuretic, but the plant American Eclectics harvested in the interior US was almost certainly U. gracilis, not U. dioica, despite being labeled with the Linnaean binomial. Felter & Lloyd’s King’s American Dispensatory (1898) carries the same attributional ambiguity. [Well-documented, with the caveat that species labels are misleading for American 19th-century sources.]
Military and wartime use. German textile use of nettle fiber during WWI (1915–18) when cotton was blockaded is well-documented [Grieve 1931]. WWII UK extraction of nettle chlorophyll for medical dyes is widely reported in secondary sources but I could not locate primary archival evidence in the time budget of this project [Gap flagged, treat as Traditionally supported rather than Well-documented pending Imperial War Museum or Kew archival confirmation].
Colonial spread and mixing. European settler agriculture carried U. dioica subsp. dioica across the Atlantic and established it widely in eastern North America from the 17th century onward; native U. gracilis was already there. The two taxa now co-occur in parts of the eastern US and Canada, and older herbarium records frequently lump them under U. dioica. Attribution of historical medicinal, fiber, and food uses in North America requires asking which nettle, the native gracilis whose uses belong to Indigenous knowledge traditions, or the introduced dioica whose uses came with European settler herbals.
Modern revival. Late-20th- and early-21st-century re-appraisal of nettle spans clinical herbalism (BPH clinical trials beginning with Vontobel 1985; Safarinejad 2005, Schneider & Rübben 2004, Lopatkin 2005, see Phase II), European regulatory acceptance (EMA HMPC community herbal monographs on Urticae radix, Urticae folium, and Urticae herba), biodynamic and regenerative agricultural practice (preparation 504; French purin d’ortie regulatory saga; German and EU fiber-nettle programs including STING and Bredemann-derived clones), and a sustained revival in home-scale food, tea, and fiber practice. [Well-documented]

9.2 Folklore and symbolism
The Wild Swans (De vilde svaner, Hans Christian Andersen, 1838). Elisa weaves eleven shirts of churchyard nettles, hands blistered and silent through the work, to disenchant her brothers. Grimm’s earlier “Six Swans” (KHM 49, 1812) uses aster; Andersen specifies nettle and roots the story in suffering-as-transformation. This is the central nettle story of world literature, redemption through contact with the stinging plant, silent labor on something that burns. [Well-documented]
Nine Herbs Charm. Lacnunga manuscript, 10th–11th c. Wergulu (nettle) is the sixth of nine herbs against “flying venom” and the “onflyings”, an Anglo-Saxon medical and magical category that encompasses both airborne contagion and supernatural malediction. The charm is sung over the herbs as ointments are made. That nettle sits in the ninefold protection alongside mugwort (mucgwyrt), plantain (wegbrade), and the others tells us that pre-Christian and early-Christian English folk medicine regarded it as fundamental [Pettit 2001; Cameron 1993]. [Well-documented]
Roman urtication. Caelius Aurelianus and Pliny document flogging paralytic or numb limbs with nettle to restore warmth and sensation. The practice persists in European rheumatic tradition for two millennia and has been validated experimentally: Randall et al. (2000) conducted a randomized controlled trial of topical U. dioica for base-of-thumb osteoarthritis and found significant pain reduction versus deadnettle control. The ancient flogging and the modern RCT are describing the same mechanism. [Well-documented]
Proverbs and idioms.
“Grasp the nettle” (English, 18th c., tracing to Aaron Hill’s 1753 verse: “Tender-handed stroke a nettle, / And it stings you for your pains; / Grasp it like a man of mettle, / And it soft as silk remains.”). The handling advice is botanically accurate at the leaf surface, firm pressure flattens the trichomes without breaking them, and the proverb elevates the observation to moral counsel.
“Nettle in, dock out, dock rub nettle out” (English folk charm). The dock-leaf pairing is widespread across British and Irish children’s lore. Dock (Rumex spp.) juice contains oxalic acid; the pharmacological mechanism for perceived sting relief is ambiguous (possibly placebo, possibly mild alkalinity) but the cultural pairing is deep [Opie & Opie 1959]. [Well-documented for the folk pairing; Emerging for the mechanism.]
Protective folklore.
Nettle against lightning, German Donnernessel; nettle hung in windows on thunderstorms [Marzell IV].
Nettle in byres against elf-shot for cattle, Anglo-Saxon leechdom tradition [Cockayne 1865].
Easter Monday / Green Thursday nettle flogging rituals in Slavic Central Europe (Carpathian villages; śmigus-dyngus variants) [Moszyński 1929–39].
Walpurgisnacht (April 30), nettle in Alpine windowsills against witches’ passage [Marzell IV].
Shakespeare. 1 Henry IV II.iii.10, Hotspur: “Out of this nettle, danger, we pluck this flower, safety.” The image is old enough that it felt obvious to an Elizabethan audience: danger handled becomes safety.
St. Columba / Colmcille. The Irish hagiographic tradition, Betha Colaim Chille, describes the monk subsisting on nettle broth and, when the trick is discovered, insisting on the practice. The story encodes nettle’s role as famine food and ascetic provision [Irish hagiographic sources].
Milarepa’s nettle diet. Tsangnyön Heruka’s 15th-century Life of Milarepa describes the Tibetan yogi subsisting on nettles in the Lapchi caves, his skin turning green. The story is hagiographic, not materia-medica-evidentiary, but it encodes a real Himalayan practice of nettle-as-ascetic-food and signals the plant’s prominence in high-altitude traditional diet [Tsangnyön 15th c.]. [Traditionally supported for the cultural association; Speculative for any biochemical claim.]
Heraldry. The Mallerstang / Malherbe families bear nettle as canting arms (mal herbe = “bad plant”) [Fox-Davies 1909]. The Nettleship surname and its heraldic devices carry the same device. [Well-documented]
9.3 Encoded agronomy
Each folklore element, read carefully, encodes a practical observation:
“Nettle in, dock out” encodes the pharmacological pairing of a sting-urticant and a juicy oxalate-containing leaf; whether or not the dock mechanism works, the pairing placed relief within arm’s reach of the injury, and taught generations of children to notice the two plants as ecological companions. Both grow in similar disturbed fertile ground.
Roman urtication for paralysis encodes the counter-irritant / histamine-release mechanism now validated for topical rheumatic pain [Randall et al. 2000]. Two thousand years of “flog the cold limb with nettles” turned out to describe a real pharmacological effect.
Scandinavian and Slavic spring nettle soup encodes the nutritional fact of spring-green iron, vitamin C, and protein after a winter of stored starches, measurable now in any nettle nutritional profile [see Phase II §12.1].
TCM xúnmá for wind-damp bi encodes the anti-inflammatory and diuretic pattern that Western clinical trials have since approached through Urticae folium for osteoarthritis [e.g., Randall et al. 2000; see Phase II §12 for the chemistry–tradition mapping].
Pacific NW Coast cordage for whaling harpoon lines encodes the bast fiber’s exceptional strength and rot resistance, the basis for STING-era European fiber-nettle research, arriving in the 21st century at the conclusion Nuu-chah-nulth and Kwakwaka’wakw cordage-makers reached in the practice [Turner & Efrat 1982; Turner & Bell 1973].
German Donnernessel against lightning has no known mechanism; the encoded fact may be simply the plant’s reliable presence at the farmyard edge and thus its symbolic availability, or it may encode nothing more than the anxiety of thunderstorm seasons and the human need for named rituals of protection.
The principle: folklore is not always empirically validated, and it is not always empirically vacant. Read it case by case. Where the practical observation has a clear mechanism, the folk tradition was running experiments on a timescale the laboratory cannot match.
9.4 Cultural caution
Several considerations of attribution are in order for this monograph.
On Indigenous North American knowledge. The Pacific Northwest Coast cordage, fishing-line, and whaling-line tradition; the urtication practices of Nuu-chah-nulth, Bella Coola, Kwakwaka’wakw, and Blackfoot peoples; the hemostatic, urinary, and postpartum uses of Nlaka’pamux, Ojibwe, Menominee, Cherokee, and Iroquois peoples, these belong to Urtica gracilis, not Urtica dioica, and belong to the specific nations whose ethnobotanists or community knowledge-keepers documented them. The Moerman NAEB database (Moerman 1998; naeb.brit.org) is the authoritative compiled English-language reference, but the primary source is always the community, the specific documenting ethnobotanist (Densmore, Smith, Turner, Hamel & Chiltoskey, Gunther, and others named here), and the knowledge-holders they worked with. Nothing in this profile should be read as a substitute for consultation with those communities’ contemporary knowledge-keepers when practical application is being considered.
On what can be shared openly. Documented use records in the peer-reviewed literature and in Moerman’s compilations are publicly available and may be cited. Ceremonial uses, restricted-knowledge preparations, and uses tied to specific protected sites often are not documented in those sources, when such uses exist, they are deliberately absent from the written record. Silence in the literature is not evidence of absence in practice.
On generalization. Indigenous North American uses of U. gracilis are documented across dozens of nations with distinct languages, territories, ecological contexts, and knowledge traditions. They do not constitute a single “Indigenous use” any more than European uses of U. dioica constitute a single “European use.” Every attribution in this profile is pinned to the specific nation and source.
On the American Eclectic 19th-century record. Samuel Thomson, William Cook, King’s American Dispensatory, and the rest of the Eclectic tradition wrote “Urtica dioica“ because that was the Linnaean binomial in circulation. The plant they were actually harvesting in the American interior was almost certainly U. gracilis. The convergence between “European U. dioica“ and “Indigenous U. gracilis“ use records may in part reflect the fact that a single biological entity was active across both, but the knowledge about how to use the North American plant traces substantially, and in many specific ways, to Indigenous peoples whose knowledge the Eclectic practitioners learned from, appropriated from, or paralleled [Cook 1869; Felter & Lloyd 1898; Moerman 1998]. [Important caveat]
10. Traditional Ecological Knowledge and Stewardship
Scope and ethics note. This section compiles publicly documented TEK from published ethnobotanical and ethnographic literature. It is not a substitute for consultation with contemporary knowledge-keepers, and it intentionally omits restricted-knowledge or ceremonial uses not in the public record.
10.1 Nations, peoples, communities associated with Urtica gracilis TEK (Indigenous North America)
Selected from peer-reviewed and university-press ethnobotanical sources. Each attribution names the nation, the specific source, and the general category of use. Detailed preparations and cultural contexts live in those sources and in the communities; they are not reproduced here in full.
Nlaka’pamux (Thompson) [Turner, Thompson, Thompson & York 1990]
Bella Coola (Nuxalk) [Smith 1928; Turner 1973]
Kwakwaka’wakw [Boas 1921; Turner & Bell 1973]
Nuu-chah-nulth (Nootka) [Turner & Efrat 1982; Drucker 1951]
Squamish, Halkomelem, other Central Coast Salish nations [Turner & Bell 1971; Turner 1995]
Makah, Quileute [Gunther 1945/1973]
Okanagan-Colville [Turner, Bouchard & Kennedy 1980]
Anishinaabe (Ojibwe) [Densmore 1928; Smith 1932]
Menominee [Smith 1923]
Potawatomi [Smith 1933]
Plains Cree [Leighton 1985]
Blackfoot [Hellson 1974]
Cherokee [Hamel & Chiltoskey 1975]
Iroquois [Herrick 1977; Rousseau 1945]
Diné (Navajo) [Wyman & Harris 1941; Mayes & Lacy 1989]
Lakota / Dakota [Rogers 1980; Gilmore 1919]
Secondary syntheses: Turner, Ancient Pathways, Ancestral Knowledge (2014); Kuhnlein & Turner, Traditional Plant Foods of Canadian Indigenous Peoples (1991); Moerman, Native American Ethnobotany (1998) and NAEB database (naeb.brit.org).
10.2 Stewardship methods
Across the Pacific Northwest Coast, selective harvest of specific patches for cordage, retting of stems in stream-side pits, and timing of harvest to stem maturity are documented practices [Turner 1995; Turner & Efrat 1982]. In many Interior and Boreal traditions, young-shoot harvest in spring for food is seasonally scheduled and associated with the return of migratory birds and the emergence of other spring greens [Densmore 1928; Leighton 1985]. [Well-documented]
In European contexts, managed nettle patches in hedgerow bases and byre corners represent a form of long-continued stewardship even when it is not framed as such, the plant has been kept, harvested, cut for compost, and left standing for butterflies, by generations of farm practice that accepts nettle as a resource rather than a problem.
10.3 Harvest ethics
Take only what is needed; leave mature stands for butterfly and other invertebrate production.
Avoid harvesting all plants from any one patch in a single pass.
Time harvest to plant stage and season, not to convenience.
Return residues and waste to the same patch or adjacent soil where possible.
Consider pollen-time (June) as a window for the plant’s own reproductive work, and light-touch the patch accordingly.
These are principles common across the documented traditions, stated here without claiming they originate in any one of them. [Traditionally supported]
10.4 Offerings and reciprocity
Practices vary widely across the documented traditions and are not generalizable. Specific reciprocal offerings at harvest are recorded in some Pacific Coast and Interior traditions but the specifics belong to those communities [Turner 1995; Turner & Bell 1973]. Readers wishing to adopt a reciprocal practice in their own context are encouraged to work with local Indigenous knowledge-keepers where possible, and otherwise to cultivate their own practice grounded in the principle of giving something back to the patch from which the harvest is taken, compost residues, mulch, care of the adjacent soil, protection of the clonal patch from excessive disturbance.
10.5 Processing traditions
European: drying on racks in shade, bundles tied at the stem base; retting of fiber stems in pits or slow water; fermentation for compost amendment (purin d’ortie); fresh-leaf infusion, decoction, or pot-herb preparation. [Well-documented, detail in Phase II §15–16.]
Indigenous North American: documented retting, cordage-twining, and basketry/textile uses across the Pacific Northwest Coast [Turner 1995; Turner & Bell 1973]; fresh-shoot or dried-leaf preparations in interior and eastern traditions [Densmore 1928; Hamel & Chiltoskey 1975]. Detail in Phase II §15.
Himalayan: retting for allo cloth (predominantly Girardinia diversifolia but sometimes U. dioica); soup (sisnu ko jhol); fresh-shoot greens; winter dried fodder [Manandhar 2002].
10.6 Permission and CARE principles
The CARE principles for Indigenous Data Governance (Collective benefit, Authority to control, Responsibility, Ethics; Carroll et al. 2020) apply to any contemporary use or publication of Indigenous North American TEK on U. gracilis. The specific implications for this monograph:
Collective benefit: TEK cited here is cited with credit to the documenting source, the nation, and, where possible, the specific knowledge-keepers named in those sources.
Authority to control: Contemporary knowledge-keepers and Tribal governments hold authority over living TEK. The 20th-century ethnobotanical compilations cited here represent a particular point in time and a particular documentation ethic; living knowledge is held by the communities, not by the literature.
Responsibility: Users of this profile who intend to apply Indigenous-sourced practices at scale, commercial cultivation, supplement manufacture, ethnobotanical education, are encouraged to work directly with the relevant communities under benefit-sharing arrangements.
Ethics: The profile’s cultural caution note (§9.4) and the attributional structure throughout are this monograph’s working interpretation of research ethics appropriate to the subject.
10.7 Alignment and divergence with modern ecological management
Modern regenerative agriculture, ecological restoration, and conservation biology have, in the last two decades, converged on several principles that were already present in the traditional European, Indigenous North American, and Himalayan nettle practices cited above:
That the plant is a resource, not a weed, on most sites where it dominates.
That managed disturbance (cutting, harvesting, grazing) improves the habitat value of nettle stands for butterflies and other invertebrates.
That long-continued stewardship of a patch (continuity of use rather than annual replanting) is compatible with long-term productivity.
That the fiber, food, medicine, and fertility-amendment uses of the plant reinforce each other in a way that monoculture agronomy tends to miss.
Divergences are equally real. Contemporary regulatory frameworks (pesticide registration, food-safety regulation, supplement manufacturing standards) apply to nettle commerce in ways that traditional practice did not anticipate. Large-scale commercial cultivation (fiber-nettle clones, supplement-industry root supply) raises sustainability questions at scales traditional practice did not encounter. The French purin d’ortie regulatory saga (see Phase II §18) is the paradigmatic case of a traditional practice colliding with a modern regulatory apparatus, and of that collision being partially resolved in favor of the traditional practice, an outcome that is not guaranteed and that required decade-long advocacy.
Phase II — The Plant in Human and Animal Hands
11. Food, Medicine, and Human Use Traditions
11.1 Culinary use
Edible parts and stages.
Young shoots and leaves — the first 4–6 inches of new spring growth, and pre-flowering leaf tips through May and June. The universal culinary material across European, Indigenous North American, Himalayan, and East Asian traditions [Grieve 1931; Turner 1995; Manandhar 2002; Kuhnlein & Turner 1991]. [Well-documented]
Seed — late summer to early autumn, harvested from mature female inflorescences. Used as condiment, nutritive sprinkle, or traditional galactagogue and tonic preparation [Grieve 1931; Weed 1989]. [Traditionally supported]
Root — autumn or early spring; primarily medicinal rather than culinary, though it appears in fermented beverages and in traditional “nettle beer” recipes [Mabey 1972]. [Traditionally supported]
Historical and contemporary preparations.
Pot-herb. Leaves blanched 2–3 minutes in boiling water to neutralize trichomes, then treated as spinach, sautéed, added to soups, folded into pasta, dropped into risotto. Blanching reduces oxalate by roughly 40–80% depending on water volume and time [Rutto et al. 2013; Adhikari et al. 2016]. The blanching water is often retained (discarded only when reducing oxalate is a priority).
Soup. Nässelsoppa (Swedish), tsouknidόpita and tsouknida (Greek), urzici (Romanian), krapivnye shchi (Russian), sisnu ko jhol (Nepali), the same functional recipe across a continent’s worth of languages: young nettle, a fat, a grain or potato, broth. [Well-documented]
Greens in pies and tarts. Greek tsouknidόpita, Turkish böreks, Italian torta pasqualina variants, Balkan pita. The plant is almost indistinguishable from spinach in these dishes once cooked.
Fermented. Lacto-fermented nettle kraut; kimchi-style preparations; traditional nettle beer (British homebrew, 18th–20th c.) [Katz 2012; Mabey 1972]. [Traditionally supported]
Dried tea. Infusion of dried leaf, typically 1–2 teaspoons dried per cup; the Susun Weed “nourishing herbal infusion” tradition uses 1 ounce dried leaf per quart, steeped 4–8 hours, as a mineral-rich daily drink [Weed 1989]. [Anecdotal but widespread in modern Western practice]
Vinegar and salt. Infused nettle vinegar is an efficient way to extract minerals and preserve spring nettle through the year; nettle-salt blends are a contemporary herbal kitchen staple.
Seed sprinkle. Dry-roasted or raw seeds used as a nutritive condiment, high in essential fatty acids (linoleic dominant; α-linolenic secondary) [Guil-Guerrero et al. 2003]. [Well-documented]
Famine and staple status. Nettle has repeatedly carried populations through scarcity, Irish famine records; British WWII nettle-gathering campaigns; Scandinavian and Slavic peasant kitchens; Himalayan lean-season sisnu. The plant is abundant, protein-dense, easy to harvest once handling is learned, and reliably available in the spring hunger gap when stored grains run low and summer crops have not yet come in.
Flavor profile. Fresh blanched young nettle: green, clean, slightly iron-forward, with a spinach-like core note and a faint marine undertone. Fully cooked: softer, more neutral, takes salt and fat readily. Aged past flowering: increasingly grassy, increasingly astringent, with a chalky note from developing cystolith calcium carbonate. The plant’s peak culinary window is narrow, two to three weeks per patch per year in temperate latitudes, and the folk rule “don’t eat nettle after it flowers” has a real chemical basis.
Culinary rationale. The convergence of cultures on essentially the same preparation, blanch, add fat, add starch, add broth, is not coincidence. The blanching neutralizes trichomes, reduces oxalate, and preserves most of the protein and minerals. The fat improves β-carotene uptake. The starch balances the mineral density. The broth recovers whatever water-soluble nutrients the blanching lifted. Any peasant kitchen that cooked nettle for more than one generation converged on the same physics.
Food pairings. Nettle takes well to: butter, cream, olive oil, yogurt; potato, barley, rye, oat; onion, leek, garlic, wild chive; sorrel, cleavers, ground elder, young dandelion, young lambs-quarters (co-harvested spring greens); egg, cheese (feta, ricotta, fresh goat); rice, pasta, polenta; salmon, trout, white fish (regional Scandinavian and Pacific Northwest pairings); mushrooms.
11.2 Western herbal traditions
Primary actions as recorded in Western herbal corpus:
Alterative / depurative / “blood cleanser”, particularly as spring tonic
Diuretic, particularly the aerial parts; traditional irrigation therapy for urinary tract complaints
Hemostatic, nosebleed, menstrual flooding, wound bleeding
Astringent, internal and external
Anti-rheumatic, both internally and via topical urtication
Anti-allergic / anti-histaminic, modern Western reframing, supported by mechanistic data [Roschek et al. 2009]
Galactagogue, postpartum tonic and milk-increaser; the evidence base is traditional rather than clinical [McIntyre 2010]
Nutritive / restorative, the modern “nourishing herbal infusion” framing
Energetics. Culpeper places the herb “under Mars”, hot and dry. Matthew Wood reads it as a remedy for a cold, damp, stagnant tissue state with scrofulous or anemic presentations [Wood 2008]. Western herbal energetics converges on nettle as a drying, warming, mineralizing plant, an opposite polarity to cooling demulcents like marshmallow or licorice.
Tissue states. The modern physio-medical tissue-state system (Wood; Trevor Stokes) frames Urtica as corrective of:
Cold/damp stagnation (especially lymphatic and urinary)
Anemic presentations with fatigue and mineral depletion
Allergic/inflammatory reactivity with boggy mucous membrane states
Arthritic congestion with cold and damp terrain [Wood 2008; Hoffmann 2003]
Preparations [Upton 2013; ESCOP 2003; EMA HMPC 2012].
Leaf infusion — 1–2 tsp dried leaf per cup, 10–15 minutes covered; 3–4 cups daily for tonic use.
Leaf nourishing infusion — 1 oz dried leaf per quart boiling water, steeped 4–8 hours covered, strained; 1–4 cups daily [Weed 1989 tradition].
Leaf tincture — typically 1:5 dried leaf in 40–50% ethanol; 2–4 mL three times daily.
Root tincture — typically 1:3 or 1:5 dried root in 50–70% ethanol; 2–6 mL two to three times daily for BPH/LUTS support.
Root decoction — 1–2 tsp dried root per cup simmered 10–15 minutes; equivalent dosing.
Freeze-dried leaf capsules — 300–600 mg per capsule; 1–3 capsules at onset of allergic symptoms per the Mittman 1990 protocol.
Topical urtication — direct stinging of affected joint or muscle, historically 20–30 seconds, repeated on consecutive days [Randall et al. 2000].
Fresh juice — extracted juice taken by the spoonful or added to water; spring tonic preparation.
Indications across the Western tradition.
BPH (Benign Prostatic Hyperplasia)/ LUTS (Lower urinary tract symptoms) (root primary): the best-supported modern indication [Safarinejad 2005; Schneider & Rübben 2004; Lopatkin 2005; Ghorbanibirgani 2013; EMA HMPC 2012].
Allergic rhinitis (leaf, freeze-dried or fresh-dried): [Mittman 1990; Roschek et al. 2009].
Rheumatic and osteoarthritic pain (leaf or topical): [Chrubasik et al. 1997; Randall et al. 2000; Riehemann et al. 1999; Chrubasik et al. 2007; EMA HMPC 2012].
Urinary tract irrigation therapy, supportive in cystitis, mild BPH, gravel [Weiss 1988; ESCOP 2003; EMA HMPC 2012].
Eczema and atopic dermatitis: infusion internally and externally [Wood 2008; Hoffmann 2003].
Spring tonic for mineral depletion, fatigue, postpartum recovery: traditional rather than clinically trialed.
Bleeding, nosebleed, menstrual flooding, wound: [Dioscorides IV.93; Culpeper 1653; Grieve 1931].
Key historical practitioners in the Western lineage.
Dioscorides (1st c.), Pliny, Galen, the classical pharmacognostic foundation.
Hildegard of Bingen (12th c.), Macer Floridus, the medieval refraction.
Fuchs (1542), Gerard (1597), Culpeper (1653), Parkinson (1640), early modern codifiers.
Felter & Lloyd (1898 King’s American Dispensatory), American Eclectic synthesis (with U. gracilis species-clarity caveat).
Maud Grieve (1931), single most-cited modern source.
Rudolf Weiss (Lehrbuch der Phytotherapie, 1960/1988), German phytotherapy standard.
David Hoffmann, Michael Moore, Matthew Wood, Susun Weed, Rosemary Gladstar, Anne McIntyre, contemporary Western herbalism.
Roy Upton (American Herbal Pharmacopoeia 2009/2013), modern consolidated monograph.
11.3 Traditional Chinese Medicine
Plant name: 蕁麻 / 荨麻 (xúnmá); also 蜇人草 zhērén cǎo, 蠍子草 xiēzi cǎo.
TCM classification [Bencao Gangmu 1596; Zhonghua Bencao 1999 vol. 2; Quanguo Zhongcaoyao Huibian 1975/1996]:
Temperature (性): 温 (warm)
Flavor (味): 辛 (acrid), 苦 (bitter)
Toxicity: 有小毒 (slightly toxic, from the sting)
Channel entry (归经): 肝 (liver), 胃 (stomach) per some sources; others add 脾 (spleen)
Direction: variable across sources; primarily dispersing-outward and moving-through
Actions: 祛风定惊 (expels wind, calms convulsion), 消食通便 (aids digestion, frees the bowels), 解毒 (resolves toxicity).
Classical indications: wind-damp bi syndrome (风濕痺, rheumatic pain with cold and damp terrain); infantile convulsions and spasms; snake and insect bites (external); urticaria (paradoxically, using the sting to treat the itch on the principle of 以毒攻毒, “using poison to attack poison”); eczema and skin eruption (external wash); abdominal stagnation; constipation.
Formulas. Nettle is not in the headline formulary of Chinese herbal medicine, it is absent from the Shennong Bencao Jing (the Han-dynasty foundational materia medica, ~200 CE) and is not among the 500 most-used herbs in modern TCM practice as compiled by Bensky et al. [Bensky, Clavey & Stöger 2004]. Its use is largely folk and regional, particularly in southwest China (Yunnan, Sichuan, Tibet), where several Urtica species are used interchangeably, U. fissa, U. laetevirens, U. hyperborea, and U. dioica s.l. [Quanguo Zhongcaoyao Huibian]. The xunma designation in TCM is thus a genus-level category rather than a precise species indication. [Well-documented for the taxonomic breadth; Traditionally supported for actions.]
TCM-specific preparations:
Dried aerial parts in decoction, typically 3–9 g per dose.
External wash with boiled decoction for skin complaints.
Wine infusion for rheumatic bi syndrome.
Fresh-plant topical application (with the sting intact) for specific traditional indications.
11.4 Ayurveda
Each of the seven classical parameters below gets its own line, even when the line must report no verified classical attribution. Silence is data.
Rasa (taste): No verified classical Ayurvedic attribution for U. dioica. [Gap / absent]
Guna (qualities): No verified classical attribution. [Gap / absent]
Virya (potency, heating/cooling): No verified classical attribution. [Gap / absent]
Vipaka (post-digestive effect): No verified classical attribution. [Gap / absent]
Prabhava (special action): No verified classical attribution. [Gap / absent]
Dosha effects: No verified classical attribution. [Gap / absent]
Dhatu / srotas: No verified classical attribution. [Gap / absent]
Honest framing. Urtica dioica has no classical Ayurvedic locus in the foundational texts, Caraka Samhita, Sushruta Samhita, Ashtanga Hridaya, Bhavaprakasha, Bhavaprakasha Nighantu, Dhanvantari Nighantu [Chopra, Nayar & Chopra 1956; Warrier et al. 1994]. The Sanskrit word vṛścikālī (वृश्चिकाली, “scorpion-like”) appears in classical materia medica but is more reliably attributed to Tragia involucrata (a stinging Euphorbiaceae) than to Urtica [Nadkarni 1908]. The northern Himalayan Amchi traditions and the Tibetan Gyud Zhi corpus carry a developed zwa-ma pharmacognosy (Section 11.5 below); these are regional Himalayan traditions rather than classical Sanskrit shastra, and they are better treated in their own terms than as “Ayurveda” [Pasang Yonten Arya 1998]. [Well-documented, for absence at classical level.]
Contemporary Indian herbal medicine does use U. dioica in Himalayan regions, Kashmir, Himachal Pradesh, Uttarakhand, where the plant occurs naturally at 8,000–10,000 ft. Kirtikar & Basu’s Indian Medicinal Plants (1918) records astringent and diuretic uses, drawing largely on European sources. These are legitimate regional traditions; they are not classical Ayurveda.
11.5 Other traditional systems
Unani / Islamic medical tradition. Qurrāṣ / anjura. Ibn Sīnā (Avicenna), Al-Qānūn fī al-Ṭibb (c. 1025), classifies as hot and dry in the second degree; seed with honey for chest congestion and asthma; leaves on malignant ulcers; diuretic; emmenagogue [Gruner / Bakhtiar translations]. Ibn al-Bayṭār’s 13th-century Al-Jāmiʿ li-mufradāt al-adwiya wa-l-aghdhiya compiles Dioscoridean content with Arabic additions. Al-Bīrūnī’s Kitāb al-Ṣaydana records the Arabic / Persian / Sanskrit synonymy [Al-Bīrūnī 11th c.]. Modern Unani practice (Hakim Ajmal Khan, CCRUM monographs) uses nettle in joint-pain formulations and digestive tonics. [Well-documented for classical Unani; Traditionally supported for modern practice.]
Prophetic medicine (Tibb al-Nabawi): no authenticated hadith mentions nettle. Later compilations (Ibn Qayyim al-Jawziyya, Al-Ṭibb al-Nabawī) do not feature qurrāṣ. The Unani tradition’s use of nettle is Greco-Arabic derivation, not scriptural [Gap / absent].
Tibetan and Himalayan. The rGyud-bzhi (Four Tantras, 12th c., attributed to Yuthok) and the Tibetan materia medica tradition employ zwa-ma in formulations for rlung (wind) disorders, cold disease, and digestive complaints [Pasang Yonten Arya 1998; Kletter & Kriechbaum 2001]. The species complex includes U. dioica, U. hyperborea, and Girardinia diversifolia, used variously across the region. Manandhar’s Plants and People of Nepal (2002) records sisnu as food, fodder, fiber (allo cloth, primarily Girardinia), rheumatic flogging agent, and medicine across Nepali traditions. Milarepa’s 15th-century hagiographic nettle diet (Tsangnyön Heruka, Life of Milarepa) has no direct materia medica weight but culturally anchors the plant in Tibetan traditional knowledge [Traditionally supported].
European folk (not covered in 11.2).
Scandinavian: nässelsoppa (Swedish); similar in Danish (nældesuppe), Norwegian (nesleklopper), Finnish nokkoskeitto; spring-tonic framing across the region [Brøndegaard 1978–80; Høeg 1974].
Slavic: Green Thursday / Maundy Thursday nettle soup; Easter Monday light flogging rituals in Carpathian villages; Christmas Eve nettle under the tablecloth [Moszyński 1929–39].
Germanic and Alpine: Nesselbier (nettle beer); Walpurgisnacht nettle in windows; Donnernessel against lightning [Marzell IV].
Balkan and Greek Orthodox: Lenten horta tradition; tsouknidόpita, nettle as one of the core Lenten greens [Della & Hadjichambis 2006].
Romanian: urzici on Palm Sunday and Easter [Borza 1968].
British Isles (Ireland and Britain): St. Columba’s nettle diet; dock-leaf pairing; Shakespearean and folk-proverbial integration [Opie & Opie 1959; Grieve 1931].
Iberian: Font Quer’s Plantas medicinales (1962), Spanish folk tonic and rheumatic urtication.
Indigenous North American — U. gracilis lineage. The ethnobotanical record is rich and specifically attributed. Recurring use-categories across many nations include:
Hemostatic / styptic — for nosebleed, wound bleeding, postpartum hemorrhage (Nlaka’pamux, Lakota, and others) [Turner et al. 1990; Rogers 1980].
Rheumatic urtication — switches applied to cold, stiff, or painful limbs (Nuu-chah-nulth, Bella Coola, Kwakwaka’wakw, Blackfoot) [Turner & Efrat 1982; Smith 1928; Turner & Bell 1973; Hellson 1974].
Urinary / kidney infusions — diuretic, for gravel, urinary complaints (Ojibwe, Menominee, Iroquois, Cherokee) [Densmore 1928; Smith 1923; Herrick 1977; Hamel & Chiltoskey 1975].
Spring greens — cooked as pot-herb (Cherokee, Iroquois, and most documented nations) [Hamel & Chiltoskey 1975; Herrick 1977; Moerman 1998].
Cordage and fiber — fishing lines, whaling harpoon lines, nets, textiles (Pacific NW Coast specifically — Bella Coola, Kwakwaka’wakw, Nuu-chah-nulth, Makah, Squamish) [Smith 1928; Turner & Bell 1973; Turner & Efrat 1982; Turner 1995; Gunther 1945/1973].
Hair wash, eczema wash, external preparations (Squamish, Halkomelem, Kwakwaka’wakw) [Turner 1995].
Postpartum tonic and childbirth preparations (Nlaka’pamux, Plains Cree) [Turner et al. 1990; Leighton 1985].
The specific preparations, dosages, and ceremonial contexts belong to the nations whose knowledge is cited; the generalities are stated here with attribution but without flattening the distinctiveness of each tradition.
Andean and Amazonian. South American ortiga covers multiple species, Cajophora, Urtica magellanica, U. echinata, and occasionally naturalized U. dioica. Bussmann & Sharon’s Peruvian ethnobotany (J. Ethnobiol. Ethnomed. 2006) records native-Urticaceae uses that are outside this profile’s strict scope but worth cross-reference. [Traditionally supported; scope caveat.]
Sub-Saharan African: no documented tradition of U. dioica medicinal use at scale, the plant is not native and minimally naturalized in most of sub-Saharan Africa. [Absent]
Pacific Islands / Polynesia: no U. dioica tradition. Urtica ferox (New Zealand tree nettle, ongaonga) has Māori cultural associations, but it is a different species in a different ecological context and falls outside this profile.
Korean and Japanese folk. Sansai (mountain vegetable) tradition includes irakusa; rural harvest of young shoots for soup and side dishes. Documented in regional ethnobotany but not well-represented in the major medical traditions of either culture. [Traditionally supported]
11.6 Cross-cultural synthesis
Five functional claims about Urtica dioica / gracilis recur across three or more unrelated cultural traditions. Where three or more independent knowledge systems, with no shared transmission path, converge on the same use, the convergence itself is evidence. The plant has made the same impression on the same human bodies across cultures. Each of these is carried forward to Section 12.5 for chemistry-tradition mapping.
(1) Hemostatic / styptic. Five independent traditions:
Greco-Roman (Dioscorides, Pliny) — nosebleed, wound bleeding
European herbal (Gerard, Culpeper, Grieve) — nosebleed, menstrual flooding
American Eclectic (Felter & Lloyd) — hemorrhage of lungs, stomach, uterus
Indigenous North American — Nlaka’pamux, Lakota, and others — postpartum bleeding, nosebleed, wounds
Unani (Ibn Sīnā) — malignant ulcers, hemorrhage
(2) Rheumatic urtication (topical flogging with nettle). Six independent traditions:
Roman (Pliny, Caelius Aurelianus)
Pacific Northwest Coast (Nuu-chah-nulth, Bella Coola, Kwakwaka’wakw)
Blackfoot (Plains North America)
Himalayan / Tibetan (sisnu flogging, zwa-ma practices)
Slavic Easter folk flogging rituals
Contemporary Western herbal (validated experimentally by Randall et al. 2000 for base-of-thumb OA)
Six-way convergence, geographically and culturally independent, on a single counter-irritant practice. This is one of the most striking ethnobotanical convergences in temperate-plant medicine.
(3) Spring tonic / pot-herb / mineral-restorative. Five+ traditions:
Scandinavian nässelsoppa
Balkan Lenten horta and tsouknidόpita
Hildegard of Bingen’s spring phlegm-purge
Cherokee, Iroquois, and other Indigenous North American spring greens
Southwest Chinese regional spring-vegetable use
Slavic Maundy Thursday soup
(4) Diuretic for urinary / kidney complaints. Six+ traditions:
Classical Greek and Roman (Galen, Dioscorides)
Western herbal lineage (Culpeper, Weiss, Hoffmann, ESCOP, EMA)
Indigenous North American — Ojibwe, Menominee, Iroquois, Cherokee — urinary decoctions
Unani (Ibn Sīnā)
Tibetan / Himalayan
Modern European phytotherapy (EMA HMPC monograph on Urticae folium)
(5) Bast fiber for cordage, nets, textiles. Four+ continents:
European Bronze Age (Lusehøj, imported from Austrian Alps)
WWI German military textiles
Pacific Northwest Coast — whaling harpoon lines, fishing nets, basketry
Himalayan allo (predominantly Girardinia but including U. dioica)
Modern European fiber-nettle programs (STING, Bredemann)
The five convergent claims are not random. They describe a consistent plant character: a high-mineral, astringent, counter-irritant, diuretic, fiber-bearing herb that humans across continents learned to use in remarkably similar ways. The chemistry behind these convergences is where Section 12 begins.

12. Chemistry, Nutrition, and Functional Compounds
12.1 Nutritional profile
Aerial parts, young-shoot stage, unless otherwise noted. Compositional figures vary by genotype, site, season, and drying method; ranges are given where published figures vary.
Macronutrients:
Crude protein: 15–30% of dry matter across plant-parts and growth stages, with young pre-flowering shoots at the upper end (20–30%) and mature aerial parts at the lower end (15–22%) [Rutto et al. 2013; Kara 2016; Adhikari et al. 2016]. Exceptionally high for a leafy green, comparable to alfalfa hay and higher than most temperate forage species. [Well-documented]
Total fiber: NDF ~25–35%, ADF ~18–25% [Kara 2016]. Moderate.
Protein amino-acid profile: complete essential amino acids; histidine, lysine, threonine, and leucine all well-represented [Rutto et al. 2013]. Favorable compared to many temperate leaf-protein sources.
Fatty acids (seed dominant): linoleic acid majority constituent (~75% of seed oil); α-linolenic acid secondary; oleic acid minor [Guil-Guerrero et al. 2003]. Seed oil is a legitimate source of essential fatty acids though nettle is not industrially pressed for oil at scale.
Minerals (per 100 g dry aerial):
Calcium: 1,400–2,900 mg [Rutto et al. 2013; Adhikari et al. 2016]. Among the highest recorded for temperate herbs.
Iron: 10–40 mg [Rutto et al. 2013]. Exceptionally high; relevant to traditional anemia/spring-tonic uses.
Magnesium: 400–800 mg [Rutto et al. 2013].
Potassium: 1,500–3,500 mg [Rutto et al. 2013].
Phosphorus: 400–600 mg.
Trace elements: Zn, Cu, Mn, Si (high), Se (variable).
Blanching reduces water-soluble mineral retention somewhat; nonetheless, cooked nettle remains exceptionally mineral-dense. [Well-documented]
Vitamins:
Vitamin A (as β-carotene): leaf is deeply green and pigment-rich; β-carotene 3–7 mg / 100 g dry [Guil-Guerrero et al. 2003].
Vitamin C: 200–500 mg / 100 g fresh young shoots; diminishes rapidly with drying and prolonged cooking. [Well-documented]
Vitamin K: high, as expected for a leafy green.
B-complex: riboflavin, thiamine, pantothenate present in modest amounts.
Chlorophyll and carotenoids: the plant’s color itself is a compositional feature, chlorophyll content at the upper end for temperate leafy greens, with multiple carotenoids including β-carotene, lutein, and xanthophylls [Guil-Guerrero et al. 2003]. This underpins both the poultry-yolk pigmentation effect and the WWII British chlorophyll-extraction program.
Anti-nutrients:
Oxalates: 1–3% of dry weight; blanching for 2–3 minutes in abundant water reduces soluble oxalate by 40–80% [Rutto et al. 2013; Adhikari et al. 2016].
Cystoliths (calcium carbonate crystals): develop in leaves through the season; prominent in flowering and post-flowering leaves; contribute to the chalky mouthfeel of late-season nettle and reduce its culinary acceptability.
Nitrate: nettle on heavily manured ground can accumulate nitrate to moderate levels; relevant for forage context more than culinary context.
Processing implications. Blanching is the standard preparation for food use: neutralizes trichomes, reduces oxalate, softens cystoliths, preserves protein and most minerals. Drying preserves most minerals and moderate amounts of protein; reduces vitamin C substantially; preserves carotenoids reasonably well if done in shade. Fermentation (lacto-fermentation, silage) reduces oxalate further and preserves mineral content; increases bioavailability of some minerals through microbial action [traditional; Kwiatkowska et al. 2015]. Freezing after blanching preserves most nutrient content for several months.
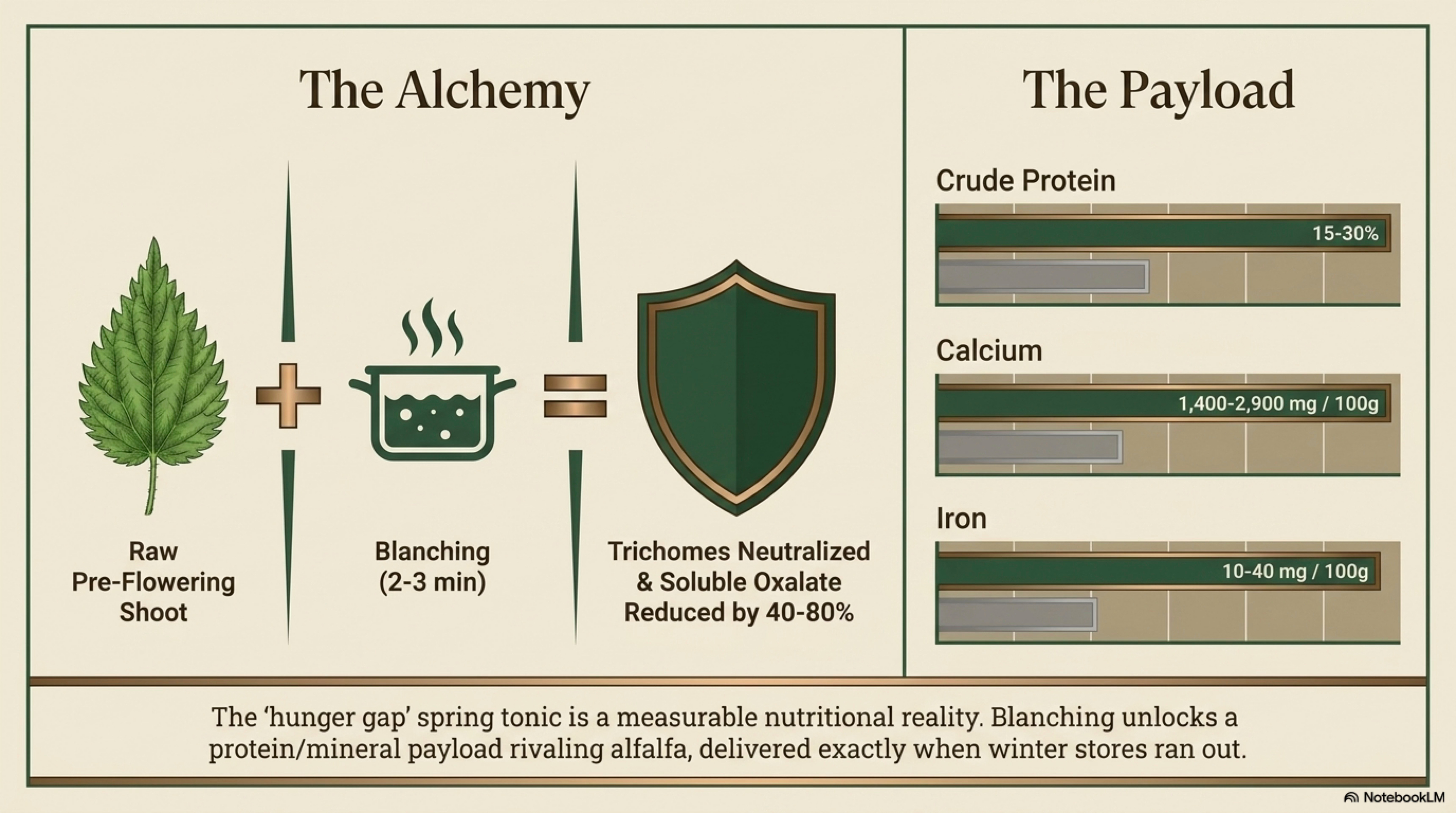
12.2 Phytochemistry
Each compound class addressed; where the answer is “no significant reports,” the absence is stated.
Flavonoids and phenolic acids [Kregiel et al. 2018; Pinelli et al. 2008; Orčić et al. 2014; Otles & Yalcin 2012; Farag et al. 2013].
Flavonol glycosides: rutin (quercetin-3-O-rutinoside), isoquercitrin, kaempferol-3-O-rutinoside, isorhamnetin-3-O-rutinoside. Quantitatively significant in leaf; less in root.
Caffeoyl-quinic acids: chlorogenic acid.
Caffeoyl-malic acid: a characteristic Urtica phenolic, linked to anti-inflammatory activity [Obertreis et al. 1996].
Other phenolic acids: caffeic, ferulic, p-coumaric.
Chemotypic variation across 43 Urtica accessions characterized by Farag et al. 2013, the largest metabolomic dataset on the genus to date. [Well-documented]
Lignans [Schöttner et al. 1997; Ganßer & Spiteller 1995].
Secoisolariciresinol, (−)-3,4-divanillyltetrahydrofuran, pinoresinol, neo-olivil, isolated from root; bind human sex hormone binding globulin (SHBG). Mechanistic foundation of the BPH hypothesis.
Lignan content largely concentrated in root rather than aerial parts, consistent with the traditional use of Urticae radix for urinary/prostatic indications. [Well-documented]
Sterols and steryl glycosides [Chaurasia & Wichtl 1987; Hirano et al. 1994].
β-sitosterol, campesterol, stigmasterol, stigmast-4-en-3-one, present in root, bioactive against prostate Na/K-ATPase.
Root-localized sterols are a second mechanistic pillar of the BPH evidence. [Well-documented]
Lectins [Peumans, De Ley & Broekaert 1984; Balzarini et al. 1992; Saul et al. 2000; Kumaki et al. 2011].
Urtica dioica agglutinin (UDA): ~8.5 kDa monomeric chitin-binding lectin, hevein-domain family, rich in glycine, cysteine, and tryptophan. N-acetylglucosamine-oligomer-specific. Localized primarily in rhizomes.
UDA has demonstrated antiviral activity in vitro against HIV, CMV, and SARS-CoV [Balzarini et al. 1992; Kumaki et al. 2011]. It is a superantigen with specific MHC interactions [Saul et al. 2000]. Recent SARS-CoV-2 spike-binding studies are emerging [Emerging].
UDA may also inhibit mycorrhizal colonization in Urtica root systems, a possible explanation for the plant’s non-mycorrhizal ecology [Emerging; hypothesis traceable to Peumans follow-ups].
[Well-documented for structural characterization and HIV/CMV activity; Emerging for SARS-CoV-2 specificity.]
Polysaccharides [Wagner et al. 1994].
Root polysaccharide fractions show anti-complement and anti-proliferative activity on prostate cells in vitro. A third mechanistic contributor to the root BPH profile. [Well-documented]
Essential oil [Gül et al. 2012].
Low total yield. GC-MS identifies carvacrol, carvone, naphthalene, (E)-anethole, and linalool among majors. Essential oil is a minor feature of the plant; not a primary medicinal vehicle.
[Well-documented for composition; Traditionally supported for practical significance.]
Stinging trichome constituents [Emmelin & Feldberg 1947; Collier & Chesher 1956; Oliver et al. 1991; Czarnetzki et al. 1990; Fu et al. 2006 (U. thunbergiana, congener)].
Histamine, acetylcholine, serotonin (5-HT) — the trichome triad. Causes the acute sting and wheal.
Leukotrienes (LTB4, LTC4-like immunoreactivity) — prolong the inflammatory response.
Oxalic acid and tartaric acid — implicated in the persistent pain phase (extrapolated from congener U. thunbergiana; primary evidence for U. dioica is less complete).
Formic acid — traditionally credited with the sting, but present at lower concentration than the amine cocktail and secondary to it [Oliver et al. 1991]. The “formic acid” narrative is a folk simplification.
[Well-documented for histamine/ACh/5-HT; Emerging for oxalate/tartrate extrapolation.]
Alkaloids: not a significant feature of Urtica dioica. Trace alkaloids may be present but are not pharmacologically meaningful [Kregiel et al. 2018]. [Well-documented absence]
Sulfur compounds: no significant sulfur-containing secondary metabolites, no glucosinolates, thiosulfinates, sulfide peptides, or sulfoxide-bearing alliums-style chemistry, have been reported for U. dioica. The plant is not a sulfur-class medicinal herb. Mineral-bound sulfur is present at expected leaf levels [Kregiel et al. 2018]. [Well-documented absence]
Saponins: present at low levels; not a primary bioactive class [Kregiel et al. 2018].
Tannins: condensed tannins are present in bark and stem tissues at modest levels; contribute to the plant’s astringent character. Hydrolyzable tannins are not a significant feature. [Traditionally supported]
Terpenes beyond essential oil: pentacyclic triterpenes (ursolic acid, oleanolic acid) reported at low levels; not a primary class [Kregiel et al. 2018].
Coumarins and polyacetylenes: not significant in U. dioica.
12.3 Functional relevance
Anti-inflammatory — strong evidence: NF-κB inhibition [Riehemann et al. 1999], caffeoyl-malic acid activity [Obertreis et al. 1996], clinical OA benefit [Randall et al. 2000; Chrubasik et al. 1997]. [Well-documented]
Antioxidant — flavonoid and phenolic acid profile supports robust in vitro antioxidant capacity [Gülçin et al. 2004; Orčić et al. 2014]. Clinical translation modest.
Antimicrobial and antiviral — UDA lectin activity against HIV, CMV, SARS-CoV in vitro [Balzarini 1992; Kumaki 2011]; modest antimicrobial activity of aerial extracts [Gülçin et al. 2004]. [Well-documented for UDA antiviral]
Nervous system / anti-allergic — leaf extract H1 antagonism, mast-cell tryptase inhibition, PGD2 synthase inhibition [Roschek et al. 2009]. Clinical support (modest) from Mittman 1990 and Bakhshaee 2017.
Digestive — modest. Traditional stomachic and mild laxative; no major clinical evidence.
Endocrine — SHBG binding by root lignans [Schöttner et al. 1997]; aromatase inhibition in vitro (minor). Clinically, the well-characterized endocrine effect is testosterone/DHT regulation in BPH context.
Immune — UDA is an immune-modulating lectin and superantigen [Saul et al. 2000]; anti-inflammatory profile supports immune-modulatory framing.
Microbiome — no specific published studies on human microbiome effects of nettle consumption; an open frontier. [Gap flagged]
Tissue-specific — prostate. Root extract effects on Na/K-ATPase in prostate [Hirano 1994], SHBG lignan binding [Schöttner 1997], polysaccharide anti-proliferative activity [Wagner 1994]. The best-characterized tissue-specific action in the plant.
Wound healing — traditional hemostatic and astringent use supported by tannin content; no modern clinical wound-healing studies.
12.4 Dynamics over time
By growth stage [Biesiada et al. 2010; Bhusal et al. 2022; traditional practice].
Pre-flowering (April–early June): peak leaf protein, chlorophyll, flavonoid glycosides, vitamin C. Lowest cystolith and lignin content. The culinary window.
Flowering (June–August): stable mineral content; declining vitamin C; increasing cystoliths; emerging seed chemistry.
Post-flowering / seed set (August–September): seed oil peak; leaf mineral content maintained but cystolith-heavy leaves less palatable; root carbohydrate and lignan content increases as the plant prepares for dormancy.
Autumn (September–November): root at peak for medicinal harvest, lignan, sterol, and polysaccharide content high; aerial parts senescing.
Winter (December–February): rhizome dormancy; underground reserves at peak; second window for root harvest before spring growth mobilizes reserves.
By plant part [Otles & Yalcin 2012; Pinelli et al. 2008; Chaurasia & Wichtl 1987; Peumans et al. 1984].
Leaf: flavonoids, phenolic acids, vitamins, minerals, chlorophyll.
Stem: bast fiber (non-chemical medicinal profile), some mineral content.
Root / rhizome: lignans, sterols, polysaccharides, UDA lectin, the BPH chemistry.
Seed: linoleic-dominant fatty oil, tocopherols (modest data), lignans (trace).
The compartmentalization maps cleanly to traditional part-selection: leaf for tonic and anti-inflammatory, root for urinary/prostatic, seed for nutritive supplement.
By stress — responsive. Nitrogen fertilization increases leaf protein and flavonoid content; drought stress can increase antioxidant enzyme activity and some phenolics. [Traditionally supported; dedicated stress-response chemistry studies thin.]
Post-harvest changes. Drying preserves most mineral content, moderate protein, chlorophyll (if in shade), and most polyphenols. Prolonged or sun-drying degrades chlorophyll and vitamin C. Freezing (after blanching) preserves most compositional features for several months. Fermentation reduces oxalate and may modify some polyphenols through microbial metabolism. [Traditionally supported; detailed dry/fresh chemistry comparison thin, gap flagged.]
Best harvest stage for different goals.
Food (maximum nutrient density): pre-flowering young shoots, April–June in temperate Europe; March–May Pacific Northwest.
Tea and tincture (aerial tonic): pre-flowering, dried in shade.
Root extraction (BPH, urinary): autumn after shoot die-back, or early spring before shoot expansion.
Seed: late summer to early autumn; just before shatter.
Fiber: late summer to autumn; full stem elongation with mature bast.
Compost / fermented amendment: vegetative through flowering stages; timing less critical for microbial conversion.
Biodynamic preparation 504: full flowering.
12.5 Chemistry–tradition convergence
(1) Hemostatic / styptic — five traditions.
Most plausible compound class: tannins (condensed), astringent phenolics, plus trichome 5-HT (which, on mucous membranes, can trigger vasoconstriction).
Quantified in U. dioica? Tannins are present at modest total levels; the quantitative hemostatic mechanism in the plant specifically, partial. Modest total content does not preclude a real local hemostatic effect at the mucous-membrane or wound-surface application where the plant was traditionally used, local concentration at the application point, not systemic dose, is the mechanism of interest. No dedicated study has measured U. dioica tannin-driven platelet-surface or fibrin-surface interaction at clinically relevant surface concentrations.
Research frontier. A targeted study of nettle aerial tannin quantification combined with in vitro whole-blood hemostasis assays would test the five-culture convergence against modern pharmacology. [Frontier Hypothesis]
(2) Rheumatic urtication — six traditions.
Most plausible compound class: the trichome triad (histamine + acetylcholine + 5-HT) combined with local oxalate/tartrate irritation, producing controlled counter-irritant inflammation and subsequent mechanistic pain modulation (likely involving TRPV1 sensitization/desensitization and local cytokine shifts).
Quantified in U. dioica? Trichome chemistry yes [Oliver 1991; Collier & Chesher 1956]; clinical validation of the counter-irritant effect yes [Randall et al. 2000, positive RCT for base-of-thumb OA with topical urtication]. The mechanism of the counter-irritation (why does stinging help arthritis?) remains incompletely characterized at the molecular level but the effect is documented.
Research frontier. Modern mechanistic study, TRPV1 involvement, local cytokine dynamics, histaminergic modulation of joint nociception, would translate six cultures of urtication practice into a modern neuro-inflammatory model. [Frontier Hypothesis; Randall 2000 is the paradigm-case for this kind of traditional-to-clinical translation.]
(3) Spring tonic / pot-herb / mineral restorative — five+ traditions.
Most plausible compound class: the mineral profile itself, Fe, Ca, Mg, K, plus essential amino acids, plus β-carotene and vitamin C. Not a pharmacological mechanism but a nutritional one.
Quantified in U. dioica? Extensively [Rutto et al. 2013; Adhikari et al. 2016; Guil-Guerrero et al. 2003]. The nutritional convergence is the best-explained of the five convergences, the plant is genuinely a high-mineral, high-protein, high-pigment spring green, and cultures that harvested it in spring were responding to measurable nutritional reality.
Research frontier. Controlled trials of spring-nettle dietary inclusion for iron-deficiency anemia and for post-winter recovery in populations with limited fresh-produce access would test the traditional-use claim at clinical endpoints. [Frontier Hypothesis, low-hanging clinical translation.]
(4) Diuretic for urinary / kidney complaints — six+ traditions.
Most plausible compound class: flavonoid glycosides (quercetin, kaempferol rutinosides, classically associated with mild aquaretic effects) and phenolic acids. Secondary contributors: potassium loading, mild smooth-muscle effects.
Quantified in U. dioica? Flavonoid content yes [Pinelli 2008; Orčić 2014; Kregiel 2018]; diuretic mechanism demonstrated in animal models [Tahri et al. 2000] but human clinical diuretic trials are few and of modest quality. The EMA HMPC monograph approves the folium for urinary irrigation therapy on traditional-use grounds, not on controlled-trial evidence.
Research frontier. A modern diuretic clinical trial of Urticae folium with quantitative urine output and electrolyte profiling would test the six-culture convergence against Western-trial standards. [Frontier Hypothesis]
(5) BPH / LUTS (specific to root).
Most plausible compound class: a triad, lignans (SHBG-binding) + sterols (Na/K-ATPase inhibition) + polysaccharides (anti-complement, anti-proliferative).
Quantified in U. dioica? Extensively, all three classes well-characterized [Schöttner 1997; Hirano 1994; Wagner 1994]. Clinical evidence: four verified RCTs (Safarinejad 2005 n=620, Schneider & Rübben 2004 n=246, Lopatkin 2005 n=257, Ghorbanibirgani 2013) with consistent modest symptom-improvement results.
Convergence story. This is the most complete chemistry-tradition translation in the nettle record: the root-BPH indication was not present in every traditional system (it is European and Unani but much less prominent in Indigenous North American records), yet where it was carried, the practitioners identified a compound-specific effect that modern chemistry has validated by three independent mechanisms.
(6) Bast fiber cordage — four+ continents.
Not a pharmacological convergence; a materials-science convergence on the same plant’s bast fibers. The chemistry here is the lignin/cellulose/pectin matrix of the stem. The convergence is evidence of the plant’s reliable mechanical properties across populations, a fact the modern STING fiber-nettle program has independently confirmed [STING Project 2005; Bredemann lineage; Vogl & Hartl 2003].
Chemistry signature of the plant. Reading all six convergences together, Urtica dioica / gracilis carries a consistent chemical signature: a nitrophilous plant that concentrates minerals and chlorophyll in leaf; develops flavonoid and caffeoyl-malic-acid-based anti-inflammatory and mild diuretic chemistry in aerial parts; localizes lignans, sterols, polysaccharides, and the UDA lectin in root; and deploys a histamine-acetylcholine-serotonin-oxalate trichome cocktail as a mammal-deterrent that humans across cultures learned to turn into a counter-irritant therapy. The cross-cultural convergences are not random; they describe the chemistry.
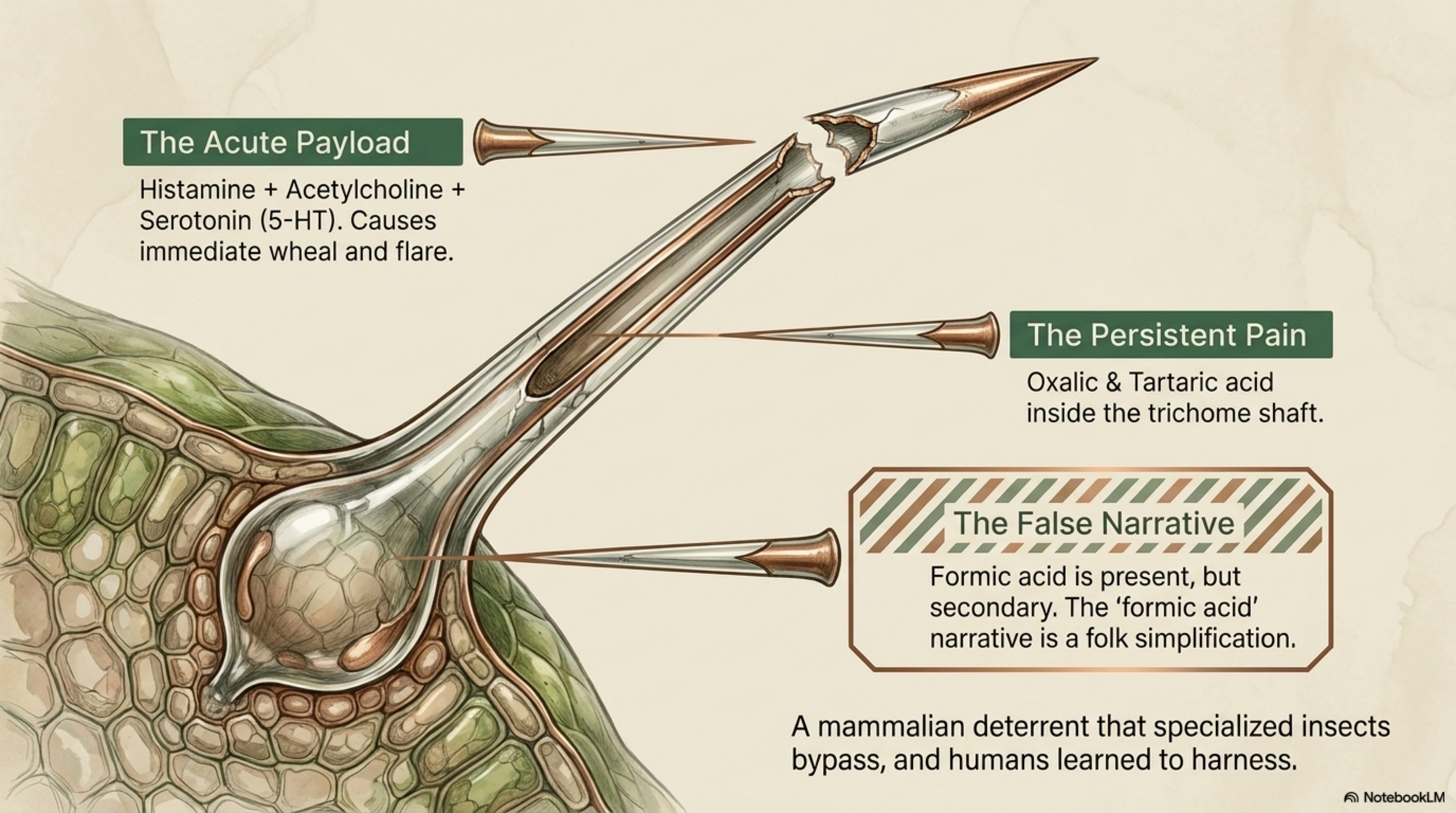
Contradictions. The clearest tension: traditional use in pregnancy. Many Western and Indigenous North American traditions used nettle in pregnancy as a nutritive tonic and as a postpartum recovery plant. Some modern herbal sources list nettle as contraindicated in pregnancy on theoretical emmenagogue grounds (echoed from Dioscorides and Culpeper seed-preparation cautions). The clinical evidence base is thin [Gap flagged]; the mechanistic basis for a pregnancy contraindication of leaf is weak; the traditional use is extensive and well-attested. This is a place where traditional use and modern caution have drifted apart without either being settled clinically.
13. Safety and Responsible Use
General profile. Urtica dioica leaf and root have a long safety record in food and medicinal use. The plant is recognized as GRAS by long history of food use in the US; the EU EMA HMPC community herbal monographs on Urticae radix, Urticae folium, and Urticae herba classify as traditional-use herbal medicinal products. Overall safety tier: A/B — food plant with a long safety record, with reasonable cautions for specific populations and specific preparations.
Toxic parts. None, in the strict toxicological sense. The stinging trichomes cause contact urticaria; the cystolith content of post-flowering leaves can cause mild gastrointestinal irritation; oxalate content warrants awareness in oxalate-sensitive individuals. No part of U. dioica carries the toxicity of, for example, comfrey (pyrrolizidine alkaloids) or foxglove (cardiac glycosides).
Safe parts and preparations.
Young pre-flowering leaves, blanched: the safest culinary form.
Dried leaf for tea, tincture, or capsule: well-tolerated at traditional doses.
Root for tincture, decoction, or capsule (BPH use): well-tolerated at traditional doses [Safarinejad 2005; Schneider 2004; Lopatkin 2005].
Seed as nutritive condiment: safe at typical dietary use.
Fresh juice: safe at teaspoon-level doses; can cause mild GI upset at larger volumes.
Preparation-dependent safety.
Fresh un-neutralized leaves: risk of contact urticaria and, if eaten raw in quantity, mild GI irritation from trichomes and cystoliths.
Blanched or cooked leaves: trichomes neutralized; oxalate reduced; safe.
Dried leaf: trichomes lose their potency; safe.
Alcohol tincture: neutralizes trichomes; safe.
Post-flowering fresh leaves: higher cystolith content; folk rule against eating nettle after flowering has a real chemical basis [Traditionally supported].
Dose-dependent concerns.
High-volume intake of fresh or lightly-cooked nettle leaf by oxalate-sensitive individuals (those with history of oxalate kidney stones) warrants moderation. Blanching reduces risk substantially [Rutto 2013; Adhikari 2016].
Very high doses of tincture (exceeding traditional 2–4 mL TID guidelines) have not been systematically studied for safety.
Pregnancy and lactation.
Leaf in food amounts: traditional across many cultures as both food and tonic in pregnancy; well-attested [Traditionally supported].
Leaf in traditional tonic doses (1–3 cups tea daily): widely used traditionally in pregnancy and lactation (galactagogue and postpartum recovery); no controlled clinical safety or efficacy data [Gap flagged].
Seed and root: historical cautions (Dioscorides; Culpeper) on seed as emmenagogue. Modern use of root in pregnancy is uncommon; avoidance is sensible on absence-of-data grounds.
Summary: leaf in food and traditional tonic amounts has strong traditional safety; seed and root warrant caution in pregnancy on precautionary grounds. [Traditionally supported for leaf; Precautionary for root/seed.]
Drug interactions [secondary aggregators — Memorial Sloan Kettering Herbs Database, Natural Medicines Comprehensive Database; primary PK studies sparse — Gap flagged].
Diuretics (Lasix, thiazides): theoretical additive effect; monitor.
Antidiabetics (insulin, sulfonylureas, metformin): theoretical additive hypoglycemic effect, potentially clinically relevant [Kianbakht 2013 showed mild HbA1c reduction in T2DM adjunctive]. Monitor blood glucose.
Antihypertensives: theoretical additive hypotensive effect [Legssyer 2002 animal data]. Monitor.
Lithium: diuretic effect could reduce lithium clearance, elevating serum lithium. Caution.
Anticoagulants and antiplatelets: nettle has high vitamin K content; theoretical interaction with warfarin dosing (nettle could reduce INR). Monitor.
CYP interactions: no significant published CYP induction or inhibition data [Gap flagged].
Allergy, dermatitis, phototoxicity.
Contact urticaria from the sting: the universal acute response; self-limiting within 30 minutes to several hours [Oliver et al. 1991]. Not an allergy per se.
True allergic reaction: rare but documented. Individuals who react atypically or severely to nettle handling should avoid handling.
Cross-reactivity: nettle pollen contributes to summer hay-fever in some individuals; this is distinct from the contact urticaria of handling.
Phototoxicity: not reported.
Oxalate, nitrate, heavy metals.
Oxalate: as discussed above. Blanch.
Nitrate: moderate on heavily manured ground; not a major concern at typical culinary or medicinal intake.
Heavy metals: nettle is a moderate accumulator of Cd, Zn, Pb, Cu on contaminated soils [Grejtovský et al. 2006]. Source selection matters: avoid harvest from roadsides, industrial brownfields, or former orchards with legacy arsenic/lead.
Fermentation concerns.
Home fermentation of nettle food products (kraut, kimchi) follows standard lacto-fermentation safety principles; salt, temperature, and anaerobic environment requirements apply as with other fermented greens.
Purin d’ortie (nettle fermented amendment) is not intended for human consumption and is regulated as a plant-protection product in the EU.
Sourcing concerns.
Wildcraft: favor clean sites; avoid nitrogen-loaded industrial margins.
Cultivated: standard organic cultivation practices apply; no documented pesticide residue concerns at typical production scales.
Supplement-market root: verify source and processing; the supplement-market supply chain for nettle root is less transparent than for some other botanicals.
Who should avoid, or use with caution.
People with history of oxalate kidney stones, moderate intake, always blanched.
People on anticoagulant therapy, monitor INR if adding significant nettle consumption.
People in pregnancy considering root or seed preparations, precautionary avoidance; leaf in traditional tonic amounts supported by long tradition.
People with specific documented nettle allergy.
People with severe renal disease, consult practitioner.
Safety tier summary: A/B. Food plant with long safety record; mainstream medicinal use with reasonable precautions; specific populations and preparations warrant modest caution; no significant toxicity.
14. Regenerative Agriculture and Land Applications
14.1 Soil and compost role
C:N and decomposition. Fresh aerial nettle: C:N roughly 10–15 [Srutek & Teckelmann 1998]; dry: 15–25 [Grime et al. 2007]. High-N litter, fast decomposition. [Well-documented]
Chop-and-drop mulch. Viable on sites where nettle is already abundant: cut pre-flowering, lay as mulch; reintegrates nitrogen, minerals, and organic matter to the local soil. Short-lived as physical mulch (decomposes in weeks on moist ground) but excellent as nutrient pulse.
Mineral contribution. Foliar mineral content (Fe, Ca, Mg, K, Si) makes nettle biomass a meaningful mineral contribution to compost systems, particularly where soils are deficient in these elements [Olsen 1921; Srutek & Teckelmann 1998]. Note the important caveat from §5.1: the high mineral content reflects fertile substrate and high plant demand, not preferential deep-soil mining. Nettle contributes minerals already present in the rootzone, not minerals pulled from depth.
Fungal vs bacterial leaning. Fast-decay, high-N litter favors bacterial decomposition pathways rather than fungal [Grime et al. 2007]. Nettle’s non-mycorrhizal ecology extends to its litter: it enriches compost biology on the bacterial-dominant side rather than the fungal-dominant side. Compost biology implications: nettle-rich compost heats up fast, processes quickly, and suits vegetable-garden use more than woody-perennial use.
Biochar synergy. No specific published studies on nettle biochar interactions; general principles apply. Pairing nettle biomass with biochar in compost likely improves biochar nutrient-charging rates.
Compost tea and extract. Aerobic compost tea with nettle as a component shows standard microbial activation and moderate nutrient solubilization; foliar application shows aphid suppression and mild disease-inhibition in field-trial literature [ITAB/INRAE]. [Emerging]
14.2 Fermentation and liquid amendment role
Purin d’ortie, the canonical European nettle fermentation amendment.
Preparation [Bertrand & Collaert 2003]:
1 kg fresh nettle (pre-flowering) per 10 L non-chlorinated water.
Fermentation vessel (plastic or wood; not metal) loosely covered.
Anaerobic-to-microaerophilic fermentation at 18–22 °C.
Stir daily; ferment 10–20 days until dark, slightly foul-smelling, no longer bubbling.
Strain; the liquid is the amendment.
Use.
Foliar spray: diluted 1:10 to 1:20 with water for pest suppression (aphids in particular), mild fungal disease suppression, and light foliar feeding.
Soil drench / root-zone amendment: diluted 1:10 for nutrient pulse and microbial stimulation; avoid undiluted application, which can burn plants.
Compost activator: dilute 1:20 added to compost to accelerate decomposition and enrich microbial community.
Microbiology [Petersen 2010s]: Lactobacillus, Bacillus, and yeast consortia dominate mature purin d’ortie. The fermentation approximates a plant-substrate lactic fermentation with aerobic Bacillus components.
KNF — Fermented Plant Juice (FPJ) adaptation. Cho Han-Kyu’s Korean Natural Farming FPJ protocol can be applied to nettle: 1:1 weight ratio plant:brown sugar, 7–10 days anaerobic ferment, strain and dilute. There is no nettle-specific KNF protocol in Cho’s published corpus; practitioners adapt the generic FPJ recipe [Gap flagged; Anecdotal for nettle-specific KNF]. The brown-sugar osmotic method produces a syrupy extract compositionally distinct from the water-based purin d’ortie.
Silage. Nettle ensiles successfully when wilted 24–48 hours and mixed with grass (30:70 nettle:grass) or supplemented with molasses; pH ~4.2; lactic acid ~6.5% DM; palatability to sheep and cattle improved vs fresh [Kwiatkowska et al. 2015; Humphries, unpublished]. [Emerging for dedicated nettle silage; Well-documented for grass-nettle mixes.]
Traditional fermentation for food. Lacto-fermented nettle kraut; nettle-based kimchi; traditional nettle beer (UK homebrew tradition) [Katz 2012; Mabey 1972]. These overlap with culinary (§11.1) and homestead (§15) treatments.
SCOBY synergies. No specific published studies on nettle in kombucha or water kefir. Practitioner reports (variable) suggest nettle leaf tea base can support healthy SCOBY growth with adjusted sugar content. [Anecdotal]
14.3 Foliar and root-zone use
Vineyard: foliar nettle extract included in some biodynamic and biological vineyard protocols for mildew suppression and micronutrient support [biodynamic literature; ITAB trials]. [Emerging]
Orchard: nettle tea foliar and soil drench recommended in Michael Phillips’s Holistic Orchard (2011); nettle patches at orchard edges as predator reservoir. [Traditionally supported]
Pasture: foliar nettle amendment not commonly practiced on pasture scale; direct nettle inclusion in forage or silage is the more common route.
Garden: purin d’ortie foliar 1:10 through the growing season; well-attested in French and German horticultural tradition. [Well-documented practice; empirical efficacy variable.]
Benefits and cautions. Modest N-P-K contribution per application (nettle slurry is not a concentrated fertilizer); real contribution is microbial activation, trace-element foliar delivery, and mild pest suppression. Undiluted application can burn foliage; dilution ratios matter. Aged slurry (>2 months) loses potency and should be refreshed.
14.4 IPM and ecosystem management
Pest-repellent / trap-crop use. Nettle hosts its own aphid (Microlophium carnosum) which does not cross to most garden vegetables, making it a functional banker plant, aphid prey populations on nettle support Coccinella (ladybirds), Aphidius parasitoid wasps, and lacewing larvae, which then disperse into adjacent crops [Hodek 1973; UK organic orchard banker-crop practice]. [Well-documented in UK organic orchard tradition]
Beneficial insect support. Butterfly host-plant value (§5.4) extends to visual and ecological value of nettle patches in mixed cropping systems. Orchards and gardens with managed nettle patches at margins typically support richer predatory arthropod communities. [Well-documented]
Disease ecology. Nettle itself is rarely seriously pest-affected at the scale that damages other crops. The plant is not a significant pathogen reservoir for common crop diseases.
Companion planting. Traditional European pairings: nettle near tomato, cucurbits (reported to improve flavor or yield, anecdotal evidence mixed); nettle near fruit trees as banker plant; nettle in herb garden margins. None of these pairings has strong controlled experimental support; most are practitioner-reported. [Anecdotal to Traditionally supported]
Push-pull systems. Not a classical push-pull component in the East African Desmodium / Striga sense; nettle’s role is more banker-plant than push-pull.
Allelopathy cautions. Nettle does not produce significant allelopathic effects on companion or successor crops [Taylor 2009]. No allelopathy-based cautions apply, a distinguishing characteristic from many other “weedy” perennials.
14.5 System fit
Garden: dedicated wild patch at the edge; source for compost activator, purin d’ortie, culinary leaf, butterfly habitat. A high-value corner on 10–50 m² of fertile ground.
Orchard: edge patches as banker plants; foliar extract for pest suppression; soil-level contribution via rhizosphere effects in long-established patches.
Vineyard: hedgerow and margin integration; biodynamic preparation 504 source (see §14.1); foliar use.
Silvopasture: native and naturalized nettle at woodland edges provides invertebrate habitat and seasonal forage-potential (wilted) for livestock.
Pasture: indicator of N/P loading; managed grazing with adequate rest can reduce dominance; otherwise accept as feature of dunged and resting zones.
Hedgerow: classic British hedgerow base-flora component; butterfly host; beneficial predator reservoir [Pollard, Hooper & Moore 1974].
Food forest: wild margin; spring green; soil conditioner; butterfly habitat.
Restoration: interim cover on nitrogen-loaded disturbed ground; phytostabilizer on moderately metal-contaminated sites [Pywell et al. 2010; Grejtovský et al. 2006].
Wild margin: the default, working with existing patches for harvest, compost, fibre, and ecological value rather than attempting eradication.
14.6 Biodynamic preparation 504
Steiner’s 1924 Agriculture Course, Lecture 5 (Koberwitz), introduces nettle as one of six compost preparations, designated 504. The protocol: dried flowering nettle inserted directly into the compost heap (no animal-organ sheath, unlike preparations 502, 503, 505, 506). The preparation is understood in biodynamic doctrine as “sensitizing” the compost to iron and sulfur flows, supporting the compost’s intelligence toward nutrient cycling [Steiner 1924; Koepf 1989; von Wistinghausen et al. 2000].
Empirical evidence:
Carpenter-Boggs, Reganold & Kennedy 2000 (RCT): biodynamic preparations collectively produced modest but statistically significant temperature-curve differences in treated compost piles; isolating preparation 504’s specific effect was not done.
Reeve et al. 2010 (WSU follow-up): no statistically significant effect of 504 alone on compost N mineralization.
Pfeiffer and earlier biodynamic case-study literature: reports of improved compost quality and agricultural outcomes; methodologically traditional rather than controlled-trial [Pfeiffer 1938/1983].
Honest framing. Biodynamic preparation 504 is a real tradition with real practitioners and a real doctrinal foundation. Controlled-trial evidence for a specific isolated effect of 504 is thin to absent. The broader question of whether biodynamic compost treatments have effects beyond conventional organic practice is contested in the agronomic literature, with some positive studies and some null studies. For a practitioner drawn to the biodynamic tradition, preparation 504 is a simple low-cost addition to the compost routine; for a practitioner demanding controlled-trial validation, the evidence is not there [Anecdotal to Emerging]. Both framings can coexist without one dismissing the other.
15. Homestead and Material Uses
15.1 Bedding
Dried nettle straw (leaf-stripped) has been used historically as livestock bedding and as human mattress filling in some Northern European rural traditions [Grieve 1931]. Not common in contemporary practice. [Traditionally supported]
15.2 Ash and lye
Wood-ash alternatives: nettle ash has a moderate potassium content and has been used traditionally in home soap-making and in horticultural potassium supplementation. Not exceptional compared to other plant ashes; reported traditionally but not a prominent contemporary use. [Traditionally supported]
15.3 Fibre, cordage, and basketry
The archaeological signal. Nettle bast fibre has been used for textiles and cordage across Eurasia and North America for at least 3,000 years [Bergfjord et al. 2012; Jacomet 2006]. The Lusehøj Bronze Age textile, imported into Bronze Age Denmark from the Austrian Alps, demonstrates that nettle cloth was sufficiently valued to move across Europe as a traded material, not merely used as a local last resort.
Pacific Northwest Coast cordage tradition. Nuu-chah-nulth whaling harpoon lines, Kwakwaka’wakw and Bella Coola fishing nets and cordage, Coast Salish and Makah twine, documented in Turner & Efrat 1982, Turner & Bell 1973, Turner 1995, Gunther 1945/1973. These uses belong to U. gracilis and to the specific coastal nations; the strength, rot-resistance, and workability of Urtica bast fibre for marine applications is a matter on which those traditions reached conclusions long before European fibre-nettle research did. [Well-documented for the tradition; attributional ethics per §10.]
Himalayan allo cloth. Predominantly Girardinia diversifolia (Himalayan giant nettle), sometimes U. dioica; distinct bast-fibre processing traditions in Nepal, Bhutan, and neighboring regions [Manandhar 2002]. Different genus, related cultural niche.
WWI German military textile program. 1915–1918: German cotton imports blocked by Allied naval blockade; nettle fibre extracted at scale for military uniform textiles [Grieve 1931]. Tens of thousands of hectares of nettle were cultivated and wild-harvested during this period. Post-war, the textile infrastructure largely dispersed.
Bredemann’s 20th-century German fibre-nettle program [see §2 of the evidence file]. 1950s–1970s breeding work, notably “Clone 13”, maintained as germplasm at Julius Kühn-Institut. Fibre content up to 16% of dry stem in selected clones [Vogl & Hartl 2003].
STING project (2002–2005). EU FP5 sustainable fibre nettle research, coordinator De Montfort University [STING Project CORDIS records]. Field yields 8–12 tonnes aerial biomass DM/ha; bast fibre yield 0.6–1.5 tonnes/ha. Processing protocols for ret-and-hackle developed for small-farm scale [Edom 2005]. [Well-documented]
Nettle-vs-ramie caveat. Ramie is Boehmeria nivea, a different genus in the same family (Urticaceae). “Nettle cloth” in historical and ethnographic literature is often ramie. Optical microscopy distinguishes them; many historical claims about “nettle fibre” conflate the two. Verify before citing any specific claim [Bergfjord & Holst 2010]. [Well-documented]
Modern small-farm practice. Retting 10–14 days in running or slowly-moving water; decortication yields 12–17% fibre by dry stem weight; ultimate nettle fibres are shorter (4–6 mm) than ramie (100–150 mm), affecting spinnability and textile feel [Dreyer & Müssig 2000s; Vogl & Hartl 2003]. Contemporary hand-spinning and small-scale textile practice is a niche but growing sector. [Well-documented for protocol]
15.4 Dye
Leaves with alum mordant: yellow-green to olive [Dean 2010].
Roots with iron mordant: grey to grey-green, sometimes trending brown.
Whole-plant ferments: more complex color profiles; practitioner knowledge varies.
Colour-fastness moderate; suitable for wool and linen, variable on cotton.
[Traditionally supported; dye chemistry not rigorously characterized.]
15.5 Soap, cleaning, and smoke
Nettle-ash lye for home soap-making: traditional but not prominent.
Nettle extract in herbal hair-rinse: widely practiced in European folk tradition and in contemporary natural-cosmetic formulation; commercial nettle shampoos and conditioners are a real market segment.
Nettle smoke/smudge: minimal documented tradition; not a classical smudge herb in any of the traditions surveyed for this profile. The absence is itself worth naming, where sage, sweetgrass, mugwort, juniper, cedar, and copal all carry smudge or incense roles in one or another tradition, nettle does not. Silence is data: the plant’s service has run through food, medicine, fibre, and soil rather than through smoke. [Gap / absent]
15.6 Building material
Not applicable at meaningful scale. Nettle fibre for paper and textile, yes; nettle for construction, no.
15.7 Feed-compost-bedding loop
The nettle-to-livestock-to-manure-to-compost-to-fertile-ground-to-more-nettle cycle is the plant’s most fundamental homestead integration. In a working system, nettle patches near byres and compost heaps self-reinforce: manure enrichment expands the patches; the patches supply wilted fodder, compost material, and preparation 504 / purin d’ortie inputs; the fertility cycles back through animals and compost to the ground that supports the next season’s nettle. This is not a technology; it is a land relationship, documented implicitly in European and Indigenous North American long-continued stewardship and available to any contemporary practitioner on fertile moist ground.
16. Harvest, Processing, and Preservation
16.1 Harvest protocols
Leaf for food and fresh medicine.
What: top 4–6 inches of new spring shoots; or pre-flowering leaf pairs from the top of young shoots.
Stage: pre-flowering (April–June in temperate Europe; March–May Pacific Northwest lowlands; shifted by latitude and elevation). “Don’t eat nettle after it flowers” is the enduring folk rule.
Weather: dry weather; morning harvest after dew has lifted and before midday heat drops the plant’s turgor.
Time of day: mid-morning to noon is traditional and practical; leaves at full turgor, trichomes fully extended and most brittle, chemistry at peak.
Ethical limits: take 20–30% of shoots from any patch in a single pass; never strip a patch entirely; rotate between patches; leave mature plants to flower and seed.
Leaf for drying.
Same stage and weather; hang in small bundles in shade with good air circulation; 4–10 days to full dry in most conditions.
Strip dry leaves from stems; store in airtight glass jars away from light.
Seed.
When: late summer to early autumn, when pendulous female inflorescences are heavy, brown, and beginning to lose their green edge.
Before shatter: if the seeds start dropping at the slightest shake, the window is closing.
How: snip whole female inflorescences into a paper bag; dry further on a drying rack; rub seeds free through a medium sieve.
Root.
When: autumn after shoot die-back, or very early spring before shoot expansion. These are the windows when rhizome carbohydrate and secondary-metabolite content are highest.
How: dig, wash thoroughly, chop into finger-length pieces; dry in shade or tincture fresh.
Ethics: root harvest is destructive to the local rhizome; plan patch-by-patch and allow years of recovery between major digs. Harvest from long-established patches with redundant biomass, not from new or marginal ones.
Fibre.
When: late summer to autumn, full stem elongation with mature bast fibre, before heavy winter weathering breaks the stems down.
How: cut at base; remove leaves and side branches; ret (submerge in slow water or pit) 10–14 days; decorticate; hackle; spin. See §15.3 for detail.
Biodynamic preparation 504 material.
Full flowering stage; dried intact and used in small handfuls in compost piles. Protocol per Koepf 1989 or von Wistinghausen et al. 2000.
Pre-flowering leaf for purin d’ortie.
Pre-flowering, abundant, vigorous growth, the same material used for culinary and fresh medicinal purposes works well for fermentation. Volume matters (1 kg fresh per 10 L water).
16.2 Quality by sense
The body’s instruments are older than the lab’s.
Smell, peak aroma.
Young nettle at full turgor, crushed between finger and thumb on an April morning when the dew has lifted and the sun has reached the patch, smells of clean green iodine. There is a marine undertone under the cut-grass note, the algal or kelp-like thread that some harvesters notice and others don’t, and that fades within an hour of cutting. If the smell has already gone hay-sweet, the harvest window on that patch has closed. If the smell is sharp but somehow thin, the patch is either drought-stressed or recently rain-chilled. If the smell is rich but carries a faintly fermented edge, leaves have been damaged and are metabolizing sugars: worth harvesting but not for the longest storage.
Dried leaf kept well: a hay-like sweetness with the mineral note underneath. A slightly chlorinated smell from over-dried leaf. A dusty staleness from leaf stored too long or in light: time to compost.
Taste.
A fresh young leaf, carefully blanched three seconds to neutralize the trichomes, put on the tongue: clean green, slightly iron-forward, spinach-adjacent but finer. A post-flowering leaf tried the same way: more astringent, drier mouthfeel, with a chalky note from cystoliths. A leaf from a drought-stressed patch: more intense, almost peppery. A leaf from a nitrogen-glutted patch (rank cow-camp, dung-heap edge): more robust, thicker texture, less delicate flavor.
Tea from dried leaf: green-hay forward, with the mineral note giving the brew its characteristic “body.” A weak green color suggests light damage during drying or old stock. A deep olive-green color on a 10-minute steep is what good dried leaf gives.
Touch.
Young shoot in the hand, gripped firmly (not tenderly): the trichomes flatten; the sting is mostly absorbed by fabric or calloused skin; the stem is hollow-soft at the top, fibrous at the base. An ungloved hand learning to harvest will learn quickly how to pinch the shoot below the first leaf pair and how to strip the leaves pad-down into a basket.
Stem past flowering: the bast fibre has begun to develop. The stem bends before it breaks; the bark peels in long strips. This is the signal for fibre harvest, if the stem snaps cleanly at a node, the fibre is still immature or already past peak.
Root, freshly dug: yellow cortex, slightly rubbery, smell faintly of turnip and damp humus. A woody, fibrous root from a long-established patch; a soft, flexible root from a younger one.
Colour.
Young shoot: a soft green at the base deepening to an almost bronze-tinged green at the tip, anthocyanin-rich from spring cold stress. The darker the tip, the stronger the folk preference for tonic use, some practitioners will harvest only the darkest-tipped shoots they can find.
Mature pre-flowering leaf: a rich even green with a slightly glaucous sheen on the upper surface. Post-flowering leaf: a slightly yellowed or grey-green edge to the otherwise even green, with cystolith dots becoming visible on close inspection.
Dried leaf: dark green-grey if dried in shade and stored well; olive-brown to yellow-brown if sun-damaged or aged; black if overheated in drying.
Sound.
A mature patch in late summer, with female inflorescences fully loaded, rustles against itself in a light wind with a sound like dry paper-crickets. At peak shatter, a sudden sharp wind will release an audible rain of achenes onto the litter, the seed-harvest window is a few days past its optimum.
The explosive stamen dehiscence of male flowers in warm still June weather, a pollen puff visible in sunlight, is quiet but not silent if you lean close.
Fresh stems, cut at the base, squeak faintly against each other in the basket on a dry day. Fibrous late-summer stems do not.
Signs of high quality.
Uniform even-green leaf colour, no yellowing or brown spots.
Clean fresh smell, no mustiness or fermentation notes.
Firm stems with intact trichomes visible when leaf held to light.
Dry leaves crisp and brittle, not rubbery; dark green with mineral fragrance.
Signs of poor quality.
Yellow, brown, or dusty-looking leaves.
Musty, moldy, or flat smell.
Soft or limp stems (water damage or post-harvest wilting).
Dried leaves that bend rather than snap (moisture damage).
Stock older than 12 months stored in clear glass or in light.
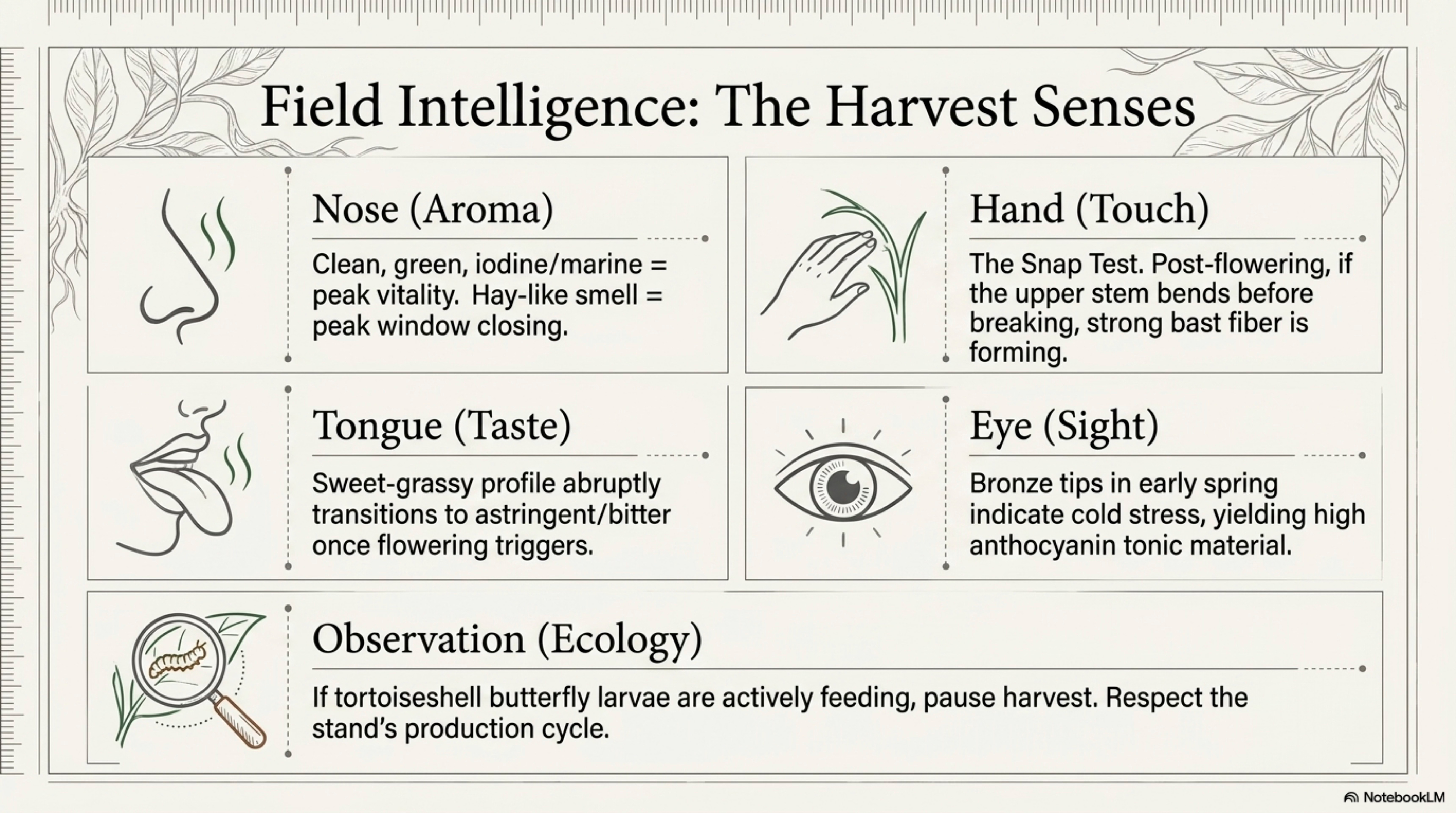
16.3 Processing and preservation
Fresh storage.
Fresh young nettle in a plastic bag with a dry paper towel: 3–5 days in refrigerator.
Longer: blanch and freeze (see below).
Drying.
Shade-dry; bundles of 5–15 stems tied at base, hung upside-down in well-ventilated shade for 4–10 days.
Finish on a drying rack at warm room temperature if needed.
Strip leaves from stems; store in airtight glass jars away from light.
Shelf life properly stored: 12–18 months at acceptable potency; beyond that, gradual potency loss.
Blanching and freezing.
Blanch 2–3 minutes in abundant boiling water; shock in ice water; squeeze gently; portion and freeze in airtight bags.
Shelf life: 6–12 months.
Retains most nutritional and culinary quality.
Fermentation.
Lacto-ferment as kraut-style preparation with 2% salt by weight; room temperature 7–14 days; refrigerate and eat within 2 months.
Purin d’ortie (not for human consumption; see §14.2).
Traditional nettle beer: specific recipes [Mabey 1972].
Tincturing.
Fresh leaf 1:2 in 95% ethanol, or dried leaf 1:5 in 40–50% ethanol; 2 weeks macerate; strain; dark glass.
Fresh root 1:2 in 95% ethanol, or dried root 1:5 in 50–70% ethanol; same protocol.
Shelf life: years.
Oil infusion.
Dried leaf in olive or sunflower oil; 2-week macerate at moderate heat (40 °C water bath) or 6-week macerate at room temperature; strain.
Used topically for rheumatic and skin preparations.
Salting and smoking.
Nettle salt: dried nettle ground with sea salt, 1:1 to 1:3 ratio; stable indefinitely; culinary staple in some modern herbal kitchens.
Smoking: not a common preservation method for nettle.
Vinegar infusion.
Fresh leaf in raw apple cider vinegar; 4–6 week macerate; strain.
A mineral-rich condiment vinegar; the vinegar also preserves the nettle for extended storage.
Residue-loop use.
Strained solids from purin d’ortie to compost.
Strained solids from tincture to compost or to secondary poultice use.
Blanching water from food preparation back to the patch (if abundant mineral-rich enough to matter), to the compost, or to livestock drink (diluted).
Stems left after leaf-stripping: retting for fibre, or direct composting.
17. Economics and Practical Value
17.1 Replacement value for farm inputs
Compost activator: nettle biomass replaces commercial compost activator products; zero marginal cost where nettle is abundant.
Purin d’ortie replaces: commercial foliar fertilizer, mild pesticide (aphid suppression), microbial activator. Retail equivalents cost €8–15/L [French market data]; home-made cost is essentially zero plus time.
Livestock forage: wilted / dried nettle as fodder supplement displaces purchased alfalfa meal or mineral supplement; on farms where nettle is abundant, savings can be material.
Biodynamic preparation 504: dried flowering nettle replaces purchased preparation from certified suppliers; cost savings modest but real for biodynamic farms.
17.2 Direct-sale value
US retail (2023–2025 benchmarks) [Anecdotal; aggregated from farmers market observations and wholesale-herbal-trade listings at Mountain Rose Herbs, Frontier Co-op, Starwest Botanicals, and allied small-scale outlets; no USDA AMS series for this crop]:
Fresh spring nettle at farmers markets: $12–20/lb retail.
Dried leaf: $30–60/lb at small-scale herbal outlets; $12–22/lb wholesale to supplement trade.
Dried root: $40–80/lb retail; $25–45/lb wholesale.
Seed: niche; $50–100/lb where sold.
Supplement market:
Nettle leaf capsules (300–500 mg): $8–15 per 60–120 capsule bottle at retail.
Nettle root capsules (BPH segment): $12–25 per bottle; premium positioning.
Global nettle supplement segment: ~$80–120 million (2023), ~6% CAGR [industry reports, Grand View Research].
Fibre market (European):
STING project costed nettle fibre at €3–5/kg processed, above flax. Viable only in eco-niche, traceable-origin, natural-textile positioning [STING Project].
Tincture and ferment products:
Nettle tincture 30 mL retail: $12–20 in the herbal / supplement market.
Commercial purin d’ortie (French): €8–15/L.
17.3 Product development opportunities
Spring-tonic infusion blends (nettle + dandelion + cleavers + red clover).
Nettle seed condiments and trail-food products.
Regional-heritage nettle textiles (fibre nettle niche, high-end natural textile).
Fermented nettle beverages (beer, cordial, vinegar).
Commercial purin d’ortie formulations for organic-horticulture markets.
Dried leaf for tea and capsule supplement trade.
Fresh nettle at spring farmers markets (highest per-pound return, narrow window).
17.4 Agritourism and education opportunities
Spring nettle harvest workshops.
Nettle fibre processing demonstrations and short-courses.
Biodynamic compost preparation workshops (preparation 504 included).
Wild-edibles courses and forage-to-table culinary events.
17.5 Scale possibilities
Household scale: zero-cost spring tonic and medicinal supply; compost activator; pest-suppression amendment. A 10–30 m² patch suffices.
Small farm scale: 0.1–1 hectare patches can supply direct-retail fresh and dried, wholesale dried leaf or root, and internal farm inputs.
Commercial cultivation scale: fibre-nettle, herbal-trade leaf and root, seed production. Feasible per European precedents (STING, Bredemann, Vogl & Hartl 2003); North American cultivation economics are not peer-reviewed documented [Gap flagged].
17.6 Cost savings
Nettle as internal farm input (compost, amendment, forage) can displace $100–500/year of purchased inputs on small farms where nettle is abundant, depending on scale and substitution rates. Specific numbers are highly context-dependent; no rigorous case studies available [Anecdotal].
17.7 Revenue potential
On the high end, a patch managed intensively for dried leaf and root, in a regional herbal market with strong demand, can return $5,000–15,000 gross per acre in spring-leaf plus root revenue [Anecdotal; small-farm reports]. These figures are unverified by peer-reviewed budget studies; real-world economics depend on market access, labor costs, and price realization.
On the low end, nettle is typically harvested as a wild resource or as a low-intensity crop component; direct revenue is modest per acre, but internal-input value (avoided costs) and ecological value (butterflies, soil, pest suppression) are significant.
17.8 Patch-scale case-study math (illustrative)
A 50 m² managed nettle patch on fertile moist ground, harvested twice in spring for fresh and dried leaf:
Fresh leaf: ~5 kg × 2 harvests = 10 kg × $15/lb retail fresh ≈ $330 gross (if direct-sale).
Or: dried leaf equivalent ~1.5 kg × $40/lb retail ≈ $130 gross (if direct-sale dried).
Plus: compost activator, purin d’ortie raw material, biodynamic preparation 504 source, butterfly habitat, seasonal culinary supply for household.
Labor: 4–8 hours of harvest and processing across the spring window.
Net: $100–300 direct revenue or input-substitution value plus non-monetized ecological and household value.
This is a stylized illustration; real economics vary. The illustration shows that even small nettle patches carry real monetary and non-monetary value to a working homestead or small farm.
The tables and price lists above matter, but they are not where the plant’s economic story actually lives. The accounting of nettle is easy to tally and easy to underestimate in the same breath. What the numbers miss is the ledger the plant keeps with the ground, and what that ledger is worth across a long-enough horizon.
17.9 Value in resilience
The deeper economic value of nettle is its reliability and redundancy. A plant that thrives on enriched disturbed ground without amendment, that carries a full-spectrum mineral and protein profile, that produces fibre and amendment and medicine and food across the same seasonal arc, that asks nothing of irrigation or fertilizer or pest management, this is the kind of plant that a farm-economy under climate and supply-chain stress increasingly cannot afford not to work with. Nettle does not displace primary crops; it occupies the margins. But the margins, in a future of increasing volatility, are where slack systems keep themselves resilient.
18. Legal, Regulatory, and Access Notes
18.1 Harvest legality
United States: no federal restrictions on harvest of U. dioica or U. gracilis on private land with owner permission or on most public land. Specific state park and federal wilderness regulations may restrict plant harvest; verify locally. Wildcraft for commercial sale requires compliance with state business licensing; no CITES or federal-species-level restrictions apply.
Canada: similar. Provincial and territorial regulations govern harvest on Crown land; First Nations and Indigenous rights may take precedence in specific territories.
United Kingdom: harvest permitted on private land with permission; on public land per Countryside and Rights of Way Act.
EU: generally permitted; specific country and site-level variation.
18.2 Protected status
US: not listed as federally endangered, threatened, or protected. Native U. gracilis is a native-plant subject of some restoration ethics but not regulatory protection.
International: not CITES-listed; not IUCN-red-listed.
18.3 Invasive restrictions
USDA APHIS Federal Noxious Weed List: NOT listed.
US state noxious weed lists: no US state lists U. dioica or U. gracilis as noxious as of 2024.
EU EPPO Global Database: not listed as quarantine pest.
Australia and New Zealand: U. dioica is locally naturalized and controlled regionally in some contexts, but is not subject to major federal invasive-species restrictions. [Well-documented]
18.4 Labeling and medicinal claim restrictions
EU: the EMA HMPC Community Herbal Monographs on Urticae folium (EMA/HMPC/508013/2006), Urticae radix (EMA/HMPC/461160/2008), and Urticae herba (adopted separately) define approved traditional-use indications: urinary tract irrigation therapy (folium and herba); lower urinary tract symptoms associated with BPH (radix); rheumatic and arthritic supportive therapy (herba, topical urtication). Products marketed within these indications and preparation specifications may carry traditional-use claims. [Well-documented]
US: DSHEA-regulated dietary supplement environment. Structure-function claims permitted with disclaimer; no FDA-approved drug claim for nettle. GRAS status by long use.
Canada: NHP (Natural Health Product) regulations; nettle products with approved DIN-HM numbers may carry specific approved claims.
18.5 Sale restrictions
Food: nettle is food. No US federal restriction on fresh or dried sale.
Supplement: nettle supplements are regulated under DSHEA (US), NHP (Canada), and THMPD / HMPC (EU). No special restrictions beyond standard herbal-supplement requirements.
Root specifically: no distinct restriction in most jurisdictions; sold as dietary supplement.
18.6 The purin d’ortie saga, a cautionary and instructive story
France, 2006: the loi d’orientation agricole (agricultural orientation law) modifies the rural code to require a full autorisation de mise sur le marché (AMM, market authorization) for any product claiming plant-protection properties. The law’s language is broad; French regulators interpret it to cover traditional home-made plant amendments sold commercially, including purin d’ortie.
Commercial sales of nettle slurry are functionally banned. Small producers face the prospect of AMM registration costs running into hundreds of thousands of euros per product, far beyond any small producer’s capacity. A grassroots campaign mobilizes: farmers, gardeners, associations including Aspro-PNPP and the GIEL. The slogan becomes: “Ce n’est pas interdit, mais ce n’est pas autorisé”, it is not forbidden, but it is not authorized. Public demonstrations, petitions, and legal challenges follow.
2011: Decree n°2011-452 (published in the Journal Officiel 28 April 2011) creates a simplified approval category, préparations naturelles peu préoccupantes (PNPP, “natural preparations of little concern”), for traditional plant-based amendments. This is a partial and principled victory: the category exists, but specific products still need approval.
2014: Arrêté of 18 April 2014, purin d’ortie is formally authorized as a PNPP for sale in France.
2017: the European Commission, in Implementing Regulation (EU) 2017/419, approves Urtica extract as a basic substance under Article 23 of Regulation (EC) No 1107/2009, the EU-level framework for plant-protection products. Nettle extract joins a small list of basic substances (equisetum, neem oil, lecithin, etc.) recognized as useful for plant protection and not requiring full pesticide-style registration. [Well-documented]
Lessons from the saga. A traditional practice that predated the regulatory framework encountered a regulatory apparatus that was not designed to accommodate it. The practice did not change; the regulation was forced to adapt, slowly, through decade-long advocacy. The outcome (traditional practice preserved; commercial sale legally available; EU-level legitimacy) was not guaranteed and required sustained collective action from practitioners who believed the tradition was worth defending. The saga is a paradigm for the collision of traditional plant practices with modern regulatory frameworks, a collision that arises repeatedly in other contexts (KNF fermented amendments, biodynamic preparations, Indigenous traditional medicines, wildcraft commerce) and will arise again. The purin d’ortie story is a reference case for how such collisions can be resolved in favor of the practice, and of what that resolution requires.
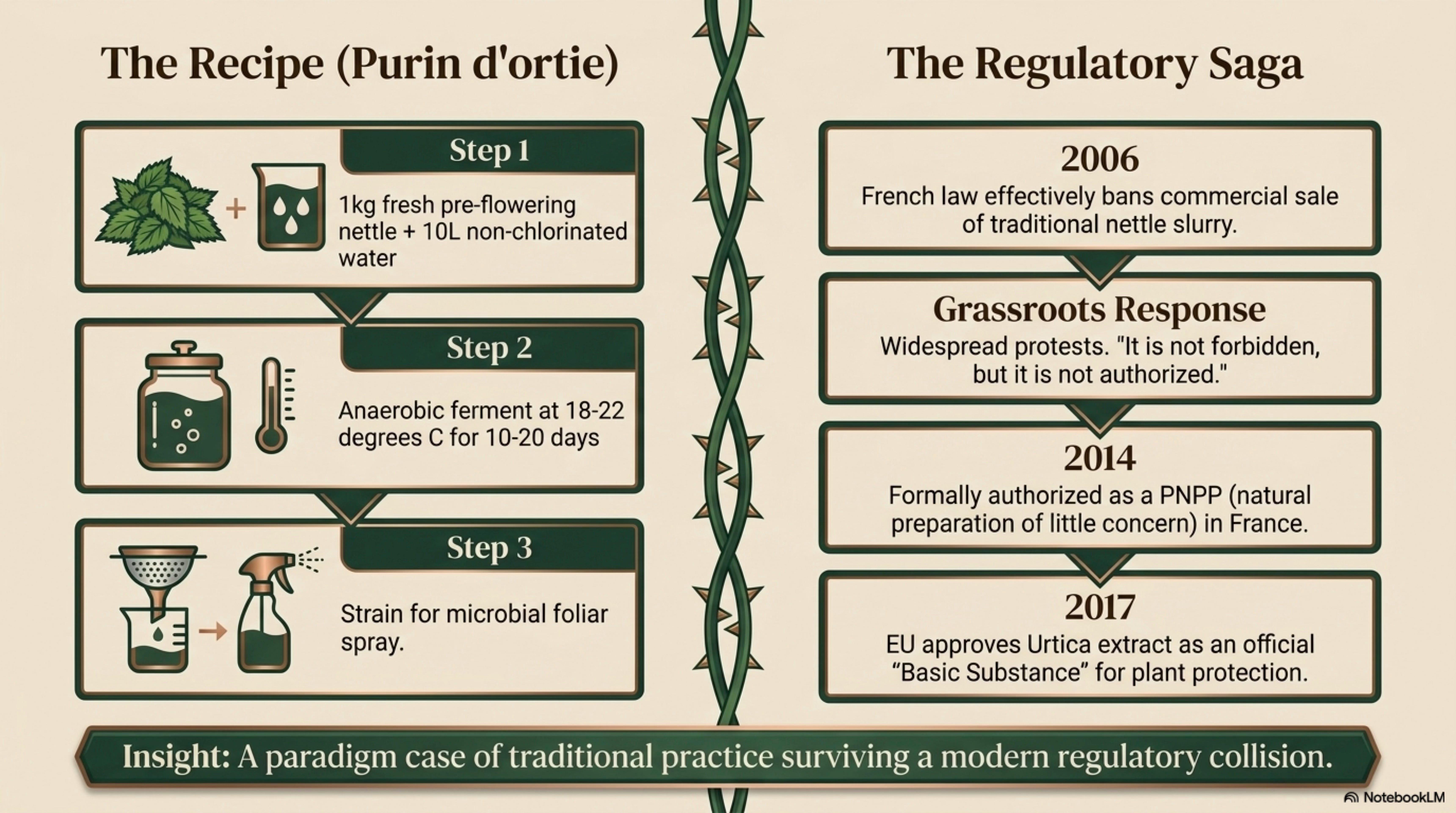
18.7 Land-access ethics
Ownership-based access: verify permission for private-land harvest.
Public-land access: verify local regulations; some jurisdictions restrict commercial wildcraft on public land.
Indigenous-territory access: harvest on Indigenous traditional territories warrants consultation with the relevant Nation. Some traditional plant-harvesting protocols belong to specific communities and cannot be unilaterally adopted by outside practitioners.
First Nations / Tribal lands: generally require tribal permission; benefit-sharing arrangements are appropriate for any commercial harvest.
18.8 Regional cautions
Some European jurisdictions regulate commercial herbal supplement sale with country-specific product-registration requirements beyond the EU-level framework.
Some US states have nuisance-weed regulations that may apply to roadside or field-margin nettle, though these are rarely enforced at scale.
No jurisdiction currently bans possession, harvest, or personal use of Urtica dioica or U. gracilis.
19. Research Frontiers and Open Questions
This section consolidates every [Gap] flag from Phases I and II into a coherent research agenda. Each frontier names the open question, why it matters, what evidence would close it, and, where applicable, which traditional observation or convergence the inquiry would honor.
19.1 Taxonomy and cytology
The gracilis question remains partly open post-split. Kew’s POWO accepts Urtica gracilis Aiton as a distinct species; USDA PLANTS and Flora of North America (1997) still treat as U. dioica subsp. gracilis. Published cytological evidence, diploid (2n=26) North American vs tetraploid (2n=52) European, predates the formal split, sits under the subspecies name in primary sources, and has not been systematically updated since POWO’s taxonomic revision. [Gap] A modern cytological and molecular phylogeographic synthesis across North American U. gracilis populations, diploid vs tetraploid distribution, sex-system frequency (monoecy vs dioecy quantified), introgression zones with introduced U. dioica subsp. dioica, western holosericea, would establish the post-split species boundary on current evidence.
19.2 Mycorrhizal ecology and root biology
U. dioica is well-documented as non- or weakly-mycorrhizal in European surveys [Harley & Harley 1987; Wang & Qiu 2006]. North American U. gracilis has not been systematically surveyed. The intriguing hypothesis, that UDA lectin, rich in the rhizome, itself inhibits mycorrhizal colonization, was raised in Peumans-era follow-up literature but has not been formally tested [Emerging]. [Gap] A controlled mycorrhizal-colonization study with paired U. dioica (European) and U. gracilis (North American) populations, including UDA-knockdown or UDA-neutralization comparisons, would establish whether the non-mycorrhizal habit is biochemically mediated.
19.3 Soil, dynamic-accumulator claim, and rhizosphere microbiome
The permaculture-lineage claim that nettle is a “dynamic accumulator”, pulling minerals from deep or impoverished soil and concentrating them, is not supported by primary experimental evidence [Taylor 2009; traces to Hamaker 1982 and Kourik 1986]. Foliar content is genuinely high on fertile sites; deep-mining is unsupported. [Gap] Rooting-depth and soil-chemistry controlled experiments, paired shallow and deep soil treatments, isotopically labeled mineral tracers, would resolve the claim decisively. A second related frontier: the rhizosphere microbiome of Urtica stands has not been systematically characterized [Gap]. The bacterial-leaning, non-mycorrhizal rhizosphere of nettle appears distinctive and likely supports the rapid nitrogen-phosphorus turnover the plant depends on, but the microbial signature is unknown.
19.4 Riparian soil-stabilization
Dense rhizome mats are widely credited with riparian bank stabilization; no peer-reviewed erosion-pin or shear-strength studies specific to Urtica were located. [Gap] Field studies on seasonally-flooded nettle stands, with shear-strength and erosion-pin comparisons against bare and other-vegetation controls, would either validate or qualify a claim that is currently on the strength of plausibility alone.
19.5 Phenology and seed-bank longevity
Seeds are “persistent” per Taylor 2009 but published longevity estimates vary widely. [Gap] A standardized soil-seed-bank burial experiment across climate zones would produce defensible longevity curves.
Phenology in U. gracilis specifically is less systematically tracked than U. dioica. [Gap] USA-NPN coverage could benefit from expanded citizen-science data, particularly in the Great Lakes, Pacific Northwest, and Boreal regions where gracilis predominates.
19.6 North American U. gracilis phytochemistry
No dedicated quantitative phytochemistry papers on U. gracilis s.s. were located. North American chemotype parity with European U. dioica is assumed, not demonstrated [Gap, major]. Lignans, sterols, UDA lectin, flavonoid profiles, and trichome constituents have all been characterized in U. dioica but not in U. gracilis. Given the diploid/tetraploid difference and the distinct evolutionary trajectory, chemotype divergence is plausible. [Gap, major; research frontier] A full chemotaxonomic comparison, ideally along the lines of Farag et al. 2013 but with U. gracilis populations included at matched developmental stages, is the single most important phytochemistry frontier for this profile. It would honor the Indigenous North American knowledge tradition by testing whether their plant is chemically what Europeans have assumed it to be.
19.7 BPH clinical evidence — no Phase III
Four verified RCTs on nettle root for BPH/LUTS (Safarinejad 2005 n=620, Schneider & Rübben 2004 n=246, Lopatkin 2005 n=257, Ghorbanibirgani 2013 n=100) show consistent modest symptom-improvement. No large multicenter Phase III trial has been conducted. A standalone Cochrane review specifically on Urtica dioica for BPH is unverified [Gap]. [Gap] A properly powered Phase III multicenter trial, with standardized Urticae radix extract, would either move BPH herbal therapy into evidence-based mainstream urology or clarify the boundary where nettle is genuinely supportive vs where pharmaceutical therapy is required.
19.8 Allergic rhinitis clinical evidence
The best-known allergic-rhinitis study (Mittman 1990) is n=98 randomized, 69 completed, 1-week duration. Bakhshaee 2017 uses root. Roschek 2009 provides mechanistic in-vitro backing. [Gap] A modern multi-week RCT of freeze-dried Urtica folium against placebo and against antihistamine standard-of-care, with quantitative symptom scoring and peripheral blood mast-cell markers, is overdue.
19.9 Rheumatic and OA clinical evidence
Randall 2000 validated topical urtication for base-of-thumb OA in a small RCT. Riehemann 1999 provides NF-κB mechanism. Obertreis 1996 supports caffeoyl-malic acid mediation. [Gap] Larger RCTs on topical urtication for knee and hand OA, combined with mechanistic investigation (TRPV1 involvement, local cytokine shifts, histamine-mediated effects on joint nociception), would translate the six-culture urtication convergence into modern neuro-inflammatory science.
19.10 Cross-cultural convergence screen — v2.1 research agenda
Convergence 1 — Hemostatic (five traditions). Tannins and trichome 5-HT are plausible mechanisms. [Frontier] Quantify condensed tannin content in U. dioica aerial extracts across growth stages and drying conditions; run whole-blood platelet aggregation and fibrin-clotting assays at clinically relevant extract concentrations. The five traditions that independently named nettle as hemostatic, Dioscorides, Culpeper, Felter & Lloyd, Ibn Sīnā, Nlaka’pamux, deserve a clean modern test.
Convergence 2 — Rheumatic urtication (six traditions). Trichome histamine + ACh + 5-HT drive acute sting; oxalate/tartrate extrapolated from U. thunbergiana likely drive persistent pain phase; counter-irritant mechanism suspected but not fully characterized. Randall 2000 validated effect for thumb-OA pain. [Frontier] TRPV1 sensitization/desensitization profile for topical nettle urtication; local cytokine dynamics (IL-6, TNF-α, IL-1β) before and after urtication; joint-nociceptor response. A research program here would translate the six-culture tradition, Roman, Pacific Northwest Coast, Himalayan, Slavic, Blackfoot, Western contemporary, into a modern counter-irritant pharmacology.
Convergence 3 — Spring tonic / mineral restorative (five+ traditions). The nutritional explanation is well-validated [Rutto 2013; Adhikari 2016]. [Frontier] Controlled trial of spring-nettle dietary inclusion (1 oz dried leaf per quart nourishing infusion, or equivalent blanched fresh) in iron-deficiency anemia populations with limited fresh-produce access. The tradition that Scandinavian, Balkan, Slavic, Cherokee, and Southwest Chinese communities converged on would be tested as a public-health intervention in populations with iron-deficiency burden.
Convergence 4 — Diuretic for urinary complaints (six+ traditions). Flavonoids + K loading plausible; animal evidence (Tahri 2000); human clinical diuretic trials are few. EMA HMPC approves Urticae folium for urinary irrigation on traditional-use grounds. [Frontier] Controlled crossover trial of Urticae folium infusion vs placebo, with quantitative 24-hour urine output, electrolyte profiling, and renal-function markers. Moves the six-culture tradition from regulatory-approved to controlled-trial-validated.
Convergence 5 — BPH root (specific to radix; narrower tradition). Three mechanistic classes (lignans, sterols, polysaccharides), four RCTs. [Frontier] Phase III multicenter RCT as per §19.7.
Convergence 6 — Bast fibre (four+ continents). Materials-science convergence rather than pharmacological. [Frontier] Standardized fibre-property comparison (ultimate fibre length, tensile strength, diameter, lignin content) between U. dioica (European fibre-nettle clones including Bredemann Clone 13) and U. gracilis populations, to test whether the North American native plant, the fibre that made Pacific Northwest Coast whaling-lines, has materials properties distinct from the European cultivar lineage.
19.11 UDA lectin — the antiviral frontier
UDA’s activity against HIV, CMV, and SARS-CoV is well-documented [Balzarini 1992; Kumaki 2011; Saul 2000 for structure]. Activity against SARS-CoV-2 is an active research area; post-2020 plant-lectin screening literature has addressed high-mannose-targeting lectins as candidates, with UDA named among them, but a definitive standalone Urtica dioica / SARS-CoV-2 peer-reviewed study was not verifiable in this research pass and should not be cited as if it were in hand [Gap pending direct verification; the broader plant-lectin screening literature is [Emerging]]. [Gap] UDA activity across a broad range of high-mannose glycan–displaying enveloped viruses (influenza, coronaviruses, filoviruses) combined with in vivo efficacy studies and translational development would move UDA from in-vitro curiosity to potential clinical asset.
19.12 Drug interactions and pharmacokinetics
Drug-interaction warnings for nettle (diuretics, antidiabetics, antihypertensives, anticoagulants, lithium) rest almost entirely on secondary aggregators [Memorial Sloan Kettering; Natural Medicines Comprehensive Database]. Primary PK studies on CYP, P-gp, or OATP interactions are essentially absent [Gap]. [Frontier] Modern PK-interaction studies with standardized nettle leaf and root extracts would replace theoretical cautions with data. This matters practically for the substantial populations using nettle alongside conventional medication for BPH, OA, diabetes, and hypertension.
19.13 Pregnancy safety evidence
Traditional use in pregnancy (leaf in food and tonic amounts) is extensive and well-attested across European and Indigenous North American traditions. Modern clinical pregnancy safety data are essentially absent [Gap]. Some herbal sources list nettle as pregnancy-contraindicated on theoretical emmenagogue grounds echoed from classical seed-preparation cautions. [Frontier] A carefully designed observational study of nettle-tea consumption in pregnancy outcomes, stratified by preparation form (leaf infusion vs tincture vs capsule vs root), would either validate the widespread traditional practice or identify preparation-specific cautions.
19.14 Zoopharmacognosy
No peer-reviewed zoopharmacognosy study of nettle-seeking behavior in wild or domestic animals has been located. Horse-owner and goat-grazier reports of deliberate animal seeking of nettle in early spring are widespread but not formally studied [Anecdotal]. [Frontier] Observational and experimental studies of livestock self-medication behavior with access to nettle, correlated with nutritional status (iron, protein, mineral balance) and with reproductive, anti-inflammatory, or antiparasitic endpoints.
19.15 North American U. gracilis cultivation economics
No peer-reviewed cultivation-economics budgets for U. gracilis in North American contexts are located [Gap]. Small-farm reports suggest $5,000–15,000 gross per acre for dried leaf and root, but these are unverified. [Frontier] Multi-site North American cultivation trial with harvest data, market realization, input costs, and labor accounting, a basic production-economics study that would support smallholder decision-making in regions where European fibre-nettle data do not directly apply.
19.16 KNF nettle-specific fermentation protocols
Cho Han-Kyu’s Korean Natural Farming corpus includes no nettle-specific FPJ/FFJ/FPE protocol [Gap]. Practitioners improvise from generic protocols. [Frontier] Systematic protocol development with microbial characterization, what does a well-executed nettle FPJ look like, chemically and microbiologically, and how does it compare to European purin d’ortie?, would make KNF-practitioner use of nettle reproducible.
19.17 Biodynamic preparation 504 — mechanism
Carpenter-Boggs 2000 and Reeve 2010 produced conflicting findings on whether biodynamic compost preparations (including 504) produce measurable compost effects beyond conventional organic practice [Emerging, contested]. [Frontier] Isolated-preparation controlled studies, with microbial community and metabolite profiling of compost with and without specific preparations, would either confirm a mechanism for the biodynamic tradition or formally establish null findings. The latter outcome would not diminish the tradition culturally but would clarify the empirical stakes.
19.18 Drying, processing, and storage chemistry
Detailed comparison of nettle chemistry (lignans, lectins, flavonoid glycosides, mineral retention) across fresh, shade-dried, sun-dried, freeze-dried, and long-stored material is thin in the primary literature [Gap]. [Frontier] A standardized storage-and-processing study would establish shelf life and preparation-dependent potency claims on evidentiary grounds.
19.19 Seed phytochemistry beyond fatty acids
Seed fatty acid profile is well-characterized [Guil-Guerrero 2003]. Seed lignans, tocopherols, and other secondary metabolites are not well-documented [Gap]. [Frontier] Comprehensive seed metabolomic profile, the traditional use of seed as galactagogue and tonic implies compounds beyond the fatty-acid story.
19.20 Microbiome effects of dietary nettle
No published studies on human microbiome effects of nettle consumption [Gap]. [Frontier] Controlled dietary intervention with gut microbiome stool-sequencing endpoints. Given nettle’s mineral and polyphenol density and the modest but real research attention to polyphenol–microbiome interactions, this is a low-hanging fruit in contemporary nutritional science.
19.21 Claims popular but weakly supported
The following claims are widely repeated in herbal and permaculture literature but are weakly supported or demonstrably anecdotal:
“Dynamic accumulator”, traced to grey literature; no primary experimental data [Anecdotal; see §19.3].
“WWII UK nettle chlorophyll extraction at industrial scale”, widely cited; primary archival evidence (Imperial War Museum, Kew archives) not located in this research pass [Traditionally supported pending archival confirmation].
“Sanskrit vṛścikālī = U. dioica“, the Sanskrit word more reliably refers to Tragia [Nadkarni 1908; Gap / misattribution flagged].
“Classical Ayurvedic materia medica includes U. dioica“, does not [§11.4; Gap].
“Nettle root-beer against scurvy” (Culpeper), historical claim worth noting but vitamin C is concentrated in leaves, not roots; root-beer ingredient specifics matter [Traditionally supported for the folk use; empirical basis for root-specific scurvy effect is unclear].
“Milarepa turned green from living on nettles”, hagiographic, not nutritional fact [Speculative; cultural significance is real].
19.22 What citizen science could help
iNaturalist re-identification of North American observations from U. dioica to U. gracilis per POWO.
Woodland Trust Nature’s Calendar and USA-NPN phenology expansion for both taxa.
Documentation of spring-harvest traditions in communities not already represented in the ethnobotanical literature (particularly post-diaspora communities in North America maintaining European nettle-soup traditions).
Home fermentation microbiology, amateur brewers and fermenters can produce observational data on purin d’ortie microbiome dynamics.
Butterfly population tracking in relation to managed vs unmanaged nettle patches.
20. Speculative, Symbolic, and Relational Layer
Every claim in this section is labeled. M = metaphor (read the plant as teacher, without empirical claim); B = belief (recorded in a tradition, without independent validation); FH = frontier hypothesis (speculative but formulated to be testable).
20.1 Doctrine of signatures and symbolic readings
The sting as teaching. [M] The plant’s first communication with a human body is a boundary. Approach wrong, the plant marks you. Approach respectfully, glove, sleeve, pinch from below the leaf, harvest in morning turgor, and the same plant offers food, medicine, fibre, amendment. The teaching encoded is older than any herbal: respect is not a substitute for knowledge; respect is knowledge’s entry requirement.
The bronze tip on young shoots. [M] Anthocyanin-rich spring tips carry a signature that European folk practice and several Indigenous traditions have independently read as iron-forward tonic material. The reading is mechanistically plausible (anthocyanin and mineral content correlate with cold-stress tissue chemistry) without being empirically validated at the dose level of folk practice. A signature reading worth taking seriously while holding it loosely.
Dioecy as signature of polarity. [M] Urtica dioica, “two-housed”, carries its male and female life on separate stems. A plant whose reproductive architecture is itself a statement about distinction. Where the North American gracilis softens this into frequent monoecy, the plant carries a different signature: not polarity but integration on a single axis. Both are the same genus, reading the same landscape differently. The signature here is about how a plant can hold both possibilities across populations.
The rhizome as colonial intelligence. [M] A nettle patch looks like a crowd. It is often a family. What appears as competition is coordinated clonal response to a single ground, with the dominant strategic decision, when to extend, when to retreat to dormancy, when to flush new shoots against a seasonal pulse, made by a network of underground organs operating on a timescale longer than any aerial shoot. If a single plant can hold decision-making distributed across meters of soil for decades, the plant’s intelligence is not metaphorical but distributed-real. The metaphor we take from this is about ourselves: the aerial life we broadcast is a partial signal of the network underneath.
The high foliar mineral profile. [M] A plant that mirrors the mineral content a human body requires, iron, calcium, magnesium, protein, on the same axis that human nutrition requires, is doing something that looks like translation. The land’s chemistry into a form the body can use. The translation is biochemically real, not metaphorical; the metaphor is the sense that the plant is offering what the place has, in the form a human body can take.
20.2 Ceremonial, dream, and story associations
Andersen’s “Wild Swans.” [B / cultural] Elisa weaves shirts from churchyard nettle, hands blistered and silent, to disenchant her brothers. The story is the canonical European literary nettle narrative: redemption is work done on something that burns, without speech, until the thing is transformed. For a culture’s mythic imagination to place nettle at that position is itself a datum, the plant sits at the intersection of suffering, silence, and transformation in a way few other plants do.
The Nine Herbs Charm / wergulu. [B] Anglo-Saxon pre-Christian and early-Christian medical magic places nettle sixth of nine plants against “flying venom.” The charm is sung over the ointment. The plant sits in the ninefold protection alongside mugwort, plantain, chamomile, wergulu, apple, chervil, fennel. Belief, specifically. The belief’s durability across ten centuries suggests something the community found true enough to preserve; the mechanism for the belief’s truth (if any) is not the point here.
Milarepa’s green skin. [B / cultural] The 15th-century Tibetan hagiography of the yogi Milarepa subsisting on nettle in the Lapchi caves, his body turning green, is not a pharmacological claim. It is a cultural narrative about ascetic transformation and the permeability of body to place. The story matters because it has been remembered, not because it happened as described.
Walpurgisnacht, Easter Monday, Green Thursday. [B] European folk flogging and apotropaic rituals on specific calendar dates, Alpine April 30, Carpathian Easter Monday, Slavic Maundy Thursday, map nettle onto the liminal moments of the seasonal year. The plant was a boundary-marker for the transition from winter into spring, from scarcity into plenty, from death into life. The ritual is the belief made concrete.
Dock-leaf pairing. [B / folk] “Nettle in, dock out, dock rub nettle out.” Children’s charm across the British Isles and Ireland. The pairing is old enough and widespread enough to suggest either a reliable pharmacological mechanism (alkaline oxalate against acidic sting; placebo by ritual relief; cold-juice vasoconstriction) or a simple co-occurrence, the two plants grow in the same habitats and the charm encoded that ecological fact into a practical remedy that works by doing something, possibly via placebo, possibly via real chemistry. The folk-belief is that it works; the mechanism is under-investigated. [B for the belief; FH for the mechanism, a simple controlled study of dock-juice effect on nettle-induced contact urticaria has not been done.]
20.3 Energetic, vibrational, and subtle-field hypotheses
Counter-irritant as energetic redistribution. [FH] The classical urtication practice, flogging a cold, stagnant, painful limb with nettle to restore warmth and sensation, has a Randall-2000 experimental validation for base-of-thumb OA and a plausible neuro-inflammatory mechanism. It also has a traditional energetic reading across six cultures: the plant “moves stuck energy,” restores circulation, “warms cold.” The energetic language and the neurological language are translating each other. [FH] The research frontier at §19.10 makes the translation testable.
UDA lectin specificity as informational selection. [FH] UDA is a lectin that recognizes high-mannose carbohydrate structures, a very specific molecular “handshake.” Its antiviral activity against enveloped viruses (HIV, CMV, SARS-CoV) reflects this specificity. The speculative reading: the plant synthesizes a molecular recognition agent in its root that is effective against pathogens humans have no evolutionary reason to share with nettle. The plant is producing, in effect, a broad-spectrum antiviral tool for reasons of its own ecology (possibly nematode defense, possibly mycorrhizal suppression), which happens to intersect with human viral pathology. Call this the “biochemical coincidence” reading, which is often how plant medicine actually operates. [FH] The broader question, whether UDA represents one instance of a general class of plant lectins with underexplored antiviral potential, is a legitimate frontier.
Biophoton and electromagnetic claims. [FH / Speculative with strong skepticism] Some strands of contemporary plant-science literature engage biophoton emission (ultra-weak photon emission from living tissue) as a signaling modality. Urtica has not been specifically studied in this framework. Claims in the energetic-herbalism literature that specific plants “resonate at specific frequencies” or “carry specific bioelectric signatures” are mostly unsupported by current mainstream physics or biochemistry. [FH] If a genuine research program on plant-cell biophoton signaling matures, nettle’s unusually clean metabolism (no major alkaloid class, no complex essential oil, consistent macronutrient profile) might make it a useful baseline model organism, but this is highly speculative and should not be confused with validated energetic medicine claims.
The silica question. [FH] Nettle has measurable silica content in its stinging-trichome tip and in its stem tissue [Thurston 1974]. Silica’s role in plant biology is well-established (structural, pathogen resistance, mineral homeostasis); its role in human health is more contested. Some traditional-use claims for nettle in bone, joint, hair, and connective-tissue conditions are framed around silica content. [FH] A controlled nutritional study of bioavailable silicon from dietary nettle, extract form, dose, absorption, connective-tissue markers, would test one of the plant’s more specific folk-medicine claims.
The “plant kingdom mirror” reading. [M / FH] Across the six cross-cultural convergences named in §11.6, hemostatic, urtication, spring tonic, diuretic, BPH, fibre, Urtica dioica appears as a plant that integrates the functions many other plants specialize in. Few plants deliver food + fibre + medicine + amendment across the same seasonal arc. The speculative reading is that nettle is, for human-temperate-latitude systems, a kind of generalist ally, a plant whose evolutionary niche happens to map onto several distinct human needs simultaneously. [M] This is not how plants think of themselves (plants do not think of themselves); it is how humans can read their relationship with this specific plant across cultures. [FH] Whether the “generalist ally” pattern is statistically distinct from other temperate herbs, whether other plants share the density of cross-cultural convergence that nettle shows, is a cross-plant comparative question a future ontology project could test by running the convergence screen across many plant profiles systematically.
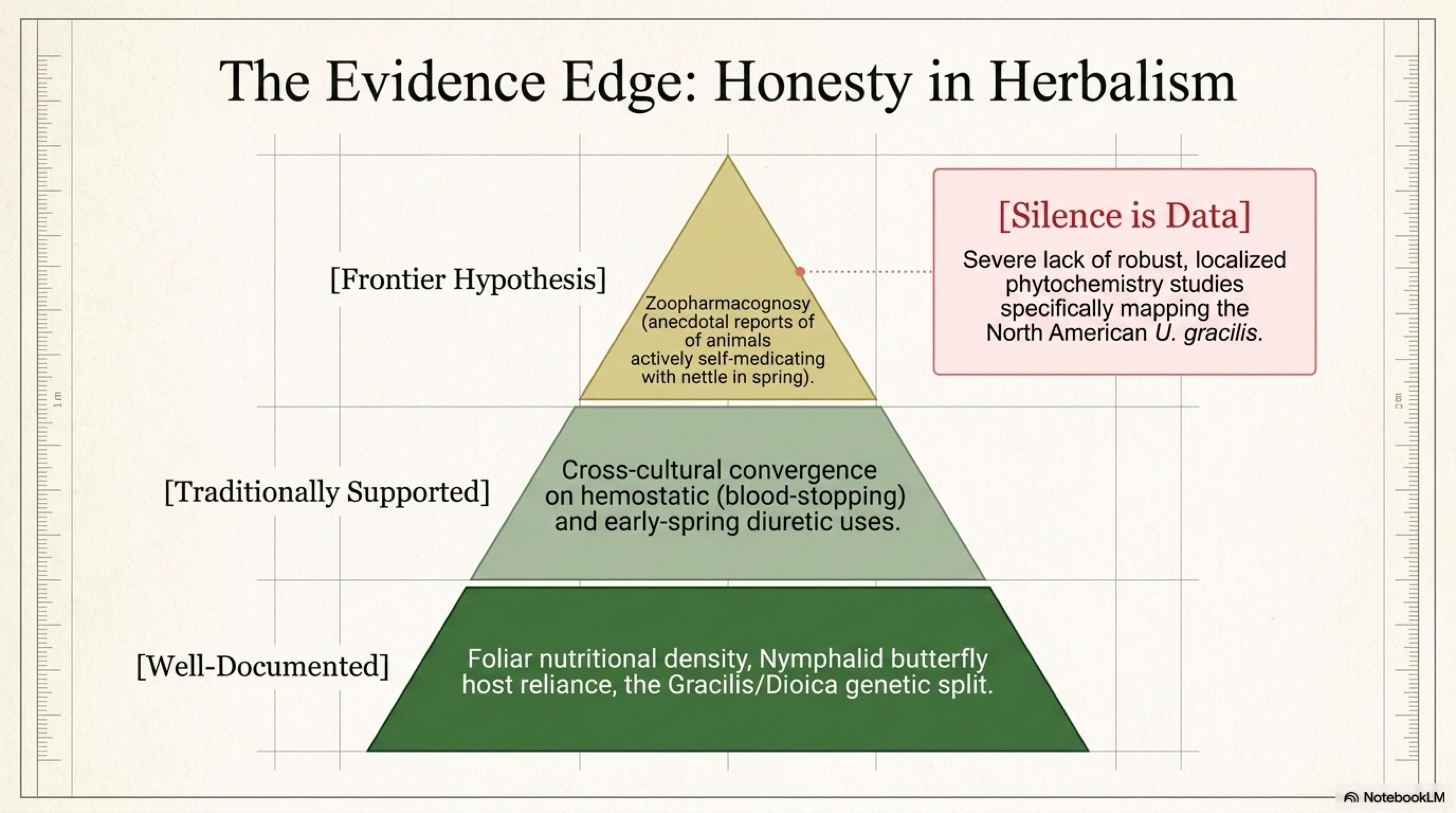
20.4 Connections to documented science
The discipline of this section, labeling every claim M, B, or FH, forces a continuous return to the empirical. Each of the speculative threads above traces back to something documented:
Trichome biology and counter-irritant pharmacology → Emmelin & Feldberg 1947, Collier & Chesher 1956, Oliver 1991, Randall 2000, Riehemann 1999.
Rhizome-localized lectin and antiviral activity → Peumans 1984, Balzarini 1992, Saul 2000, Kumaki 2011.
Foliar mineral content and nutritional restoration → Rutto 2013, Adhikari 2016.
Silica in plant and trichome structure → Thurston 1974.
Cross-cultural convergence as methodology → v2.1 ontology template; Moerman 1998; cross-referenced with Dioscorides, Ibn Sīnā, TCM and Tibetan sources.
The speculative layer is not a retreat from evidence. It is the honest labeling of where the evidence runs out, of what the traditions held as belief, of what the plant has taught metaphor to generations, and of what the testable questions are that would move the speculative into the empirical. The layer exists because a plant is more than the sum of its validated findings, and because saying so without discipline is dishonest. The MBFH labels are the discipline.
21. Sources, Confidence, and Citation Architecture
21.1 Confidence tagging system
[Well-documented] — multiple peer-reviewed sources, consistent across independent studies.
[Traditionally supported] — consistent across cultures or long-documented in practice, limited formal study.
[Emerging] — single studies, preliminary data, recent findings not yet replicated.
[Anecdotal] — field reports, practitioner observations, personal experience. Valuable but uncorroborated.
[Speculative] — hypothesis or pattern recognition not yet subjected to formal inquiry.
[Gap] — evidence does not yet exist; absence of evidence named explicitly.
Section 20 adds three further tags specific to the speculative layer:
[M] Metaphor — read the plant as teacher, without empirical claim.
[B] Belief — recorded in a tradition, without independent validation.
[FH] Frontier Hypothesis — speculative but formulated to be testable.
21.2 Source categories
The profile draws from:
Peer-reviewed primary literature — the phytochemistry, pharmacology, clinical-trial, and ecological research anchors. Taylor 2009; Safarinejad 2005; Schneider & Rübben 2004; Peumans 1984; Hirano 1994; Schöttner 1997; Rutto 2013; Bergfjord 2012; Pigott & Taylor 1964; Grejtovský 2006; Riehemann 1999; Randall 2000; Mittman 1990; Kregiel 2018 and others.
Peer-reviewed review articles and monographs — Kregiel et al 2018 Molecules; Chrubasik et al 2007 Phytomedicine; Upton 2013 Journal of Herbal Medicine; Upton (ed.) 2009 American Herbal Pharmacopoeia.
Classical and early modern herbals — Dioscorides De Materia Medica (1st c.); Pliny Naturalis Historia (1st c.); Galen (2nd c.); Ibn Sīnā Al-Qānūn fī al-Ṭibb (c. 1025); Ibn al-Bayṭār (13th c.); Hildegard of Bingen Physica (12th c.); Fuchs 1542; Gerard 1597; Parkinson 1640; Culpeper 1653.
Ethnobotanical compilations and databases — Moerman 1998 Native American Ethnobotany; NAEB database (naeb.brit.org); Kuhnlein & Turner 1991; Turner 1995; Turner & Bell 1971, 1973; Turner & Efrat 1982; Densmore 1928; Smith 1923, 1928, 1932, 1933; Hamel & Chiltoskey 1975; Hellson 1974; Wyman & Harris 1941; Leighton 1985; Rogers 1980; Manandhar 2002.
Foundational ontological and philosophical texts within Jay’s working canon — Moerman’s Native American Ethnobotany as the cornerstone Indigenous North American reference; classical herbal tradition from Dioscorides and Pliny forward; Grieve’s A Modern Herbal (1931) as the Western herbal bridge; Culpeper’s Complete Herbal (1653) for energetic/astrological tradition. Palmer, Phillips, and allied regenerative-practitioner reading on orchard integration and biodynamic tradition (Phillips 2011 The Holistic Orchard; biodynamic corpus Steiner 1924, Pfeiffer 1938, Koepf 1989, Carpenter-Boggs 2000).
Traditional medicine compendia — Bencao Gangmu (Li Shizhen 1596); Zhonghua Bencao (1999); Quanguo Zhongcaoyao Huibian (1975/1996); rGyud-bzhi (12th c.); Kirtikar & Basu 1918; Chopra 1956; Warrier 1994; Bensky et al 2004.
Regulatory and grey literature — EMA HMPC monographs (2006, 2008, 2017); ESCOP 2003; USDA PLANTS; POWO; Flora of North America; Flora of China; CABI; STING project CORDIS records; French JO (decrees 2011-452, arrêté 18 April 2014); EU Implementing Regulation 2017/419; WSSA Heap database; EPPO.
Practitioner and field-guide literature — Hoffmann, Weiss, Wood, Moore, Weed, Gladstar, McIntyre, Mabey, Edom, Phillips. Drawn on for practice and preparation detail; empirical claims back-checked to primary sources where relevant.
21.3 Citation format
Inline citations in the body of the monograph follow [Author Year] format, with fuller references in the alphabetical bibliography below (§21.4). Where a claim rests on a traditional source without a modern peer-reviewed equivalent, the tradition is cited directly (Dioscorides IV.93; Culpeper 1653 s.v. Nettles). Where a claim rests on a specific Indigenous knowledge tradition, the source is pinned to the nation, the documenting ethnobotanist, and the publication (e.g., [Turner & Efrat 1982] for Nuu-chah-nulth).
Where this profile has been unable to verify a specific citation (Cochrane review for Urtica dioica BPH; specific SARS-CoV-2 UDA papers; WWII UK chlorophyll primary archives), the citation is marked with a gap flag and the absence is named in §19 rather than papered over.
21.4 Compiled references
Alphabetical. Full citations for sources cited across Phases I–III. Where a DOI or URL is available, it is provided.
Adhikari BM, Bajracharya A, Shrestha AK. (2016). Comparison of nutritional properties of stinging nettle (Urtica dioica) flour with wheat and barley flours. Food Science & Nutrition 4(1):119–124. doi:10.1002/fsn3.259.
Al-Bīrūnī (11th c.). Kitāb al-Ṣaydana fī al-Ṭibb.
Annenkov NI. (1878). Botanicheskiĭ Slovar’. St. Petersburg.
Bakhshaee M, Mohammadpour AH, Esmaeili M, et al. (2017). Efficacy of supportive therapy of allergic rhinitis by stinging nettle (Urtica dioica) root extract. Iranian Journal of Pharmaceutical Research 16(Suppl):112–118.
Balzarini J, Neyts J, Schols D, et al. (1992). The mannose-specific plant lectins... and the Urtica dioica lectin are potent inhibitors of HIV and CMV replication in vitro. Antiviral Research 18(2):191–207. doi:10.1016/0166-3542(92)90038-7.
Bassett IJ, Crompton CW, Woodland DW. (1974). The family Urticaceae in Canada. Canadian Journal of Botany 52:503–516. doi:10.1139/b74-066.
Baytop T. (1999). Türkiye’de Bitkilerle Tedavi [Therapy with Plants in Turkey]. Nobel Tıp.
Bensky D, Clavey S, Stöger E. (2004). Chinese Herbal Medicine: Materia Medica, 3rd ed. Eastland Press.
Bergfjord C, Holst B. (2010). A procedure for identifying textile bast fibres using microscopy. Ultramicroscopy 110:1192–1197.
Bergfjord C, Mannering U, Frei KM, et al. (2012). Nettle as a distinct Bronze Age textile plant. Scientific Reports 2:664. doi:10.1038/srep00664.
Bertrand B, Collaert J-P. (2003). Purin d’ortie et compagnie. Éditions de Terran.
Biesiada A, Kucharska A, Sokół-Łętowska A, Kuś A. (2010). Effect of plantation age and harvest term on chemical composition and antioxidant activity of stinging nettle. Ecological Chemistry and Engineering A 17(9):1061–1068.
Bhusal KK, Magar SK, Thapa R, et al. (2022). Nutritional and pharmacological importance of stinging nettle (Urtica dioica L.): A review. Heliyon 8(6):e09717. doi:10.1016/j.heliyon.2022.e09717.
Bobbink R, Hicks K, Galloway J, et al. (2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity. Ecological Applications 20:30–59. doi:10.1890/08-1140.1.
Boas F. (1921). Ethnology of the Kwakiutl. 35th Annual Report, Bureau of American Ethnology.
Boufford DE. (1997). Urticaceae. In: Flora of North America Editorial Committee, eds., Flora of North America North of Mexico, Vol. 3. Oxford University Press.
Borza A. (1968). Dicţionar etnobotanic. Editura Academiei RSR.
Brøndegaard VJ. (1978–80). Folk og Flora: Dansk etnobotanik. Rosenkilde og Bagger.
CABI (2023). Urtica dioica (stinging nettle). CABI Compendium. doi:10.1079/cabicompendium.55909.
Cameron ML. (1993). Anglo-Saxon Medicine. Cambridge University Press.
Carpenter-Boggs L, Reganold JP, Kennedy AC. (2000). Effects of biodynamic preparations on compost development. Biological Agriculture & Horticulture 17:313–328.
Carroll SR, Garba I, Figueroa-Rodríguez OL, et al. (2020). The CARE Principles for Indigenous Data Governance. Data Science Journal 19:43.
Chaurasia N, Wichtl M. (1987). Sterols and steryl glycosides from Urtica dioica. Journal of Natural Products 50(5):881–885. doi:10.1021/np50053a018.
Chopra RN, Nayar SL, Chopra IC. (1956). Glossary of Indian Medicinal Plants. CSIR New Delhi.
Chrubasik JE, Roufogalis BD, Wagner H, Chrubasik S. (2007). A comprehensive review on the stinging nettle effect and efficacy profiles. Part II: Urticae radix. Phytomedicine 14(7–8):568–579. doi:10.1016/j.phymed.2007.03.014.
Chrubasik S, Enderlein W, Bauer R, Grabner W. (1997). Evidence for antirheumatic effectiveness of Herba Urticae dioicae in acute arthritis. Phytomedicine 4(2):105–108.
Cockayne O. (1865). Leechdoms, Wortcunning, and Starcraft of Early England. Rolls Series.
Collier HOJ, Chesher GB. (1956). Identification of 5-hydroxytryptamine in the sting of the nettle (Urtica dioica). British Journal of Pharmacology and Chemotherapy 11(2):186–189.
Connor HE. (1977). The Poisonous Plants in New Zealand, 2nd ed. Government Printer, Wellington.
Cook WH. (1869). Physiomedical Dispensatory. William H. Cook, Cincinnati.
Culpeper N. (1653). The English Physitian; or, an Astrologo-Physical Discourse of the Vulgar Herbs of this Nation (Complete Herbal). Peter Cole, London.
Czarnetzki BM, Thiele T, Rosenbach T. (1990). Immunoreactive leukotrienes in nettle plants (Urtica urens). International Archives of Allergy and Applied Immunology 91(1):43–46.
Daher CF, Baroody KG, Baroody GM. (2006). Effect of Urtica dioica extract intake upon blood lipid profile in the rats. Fitoterapia 77(3):183–188.
Dean J. (2010). Wild Color. Watson-Guptill.
Della A, Hadjichambis ACh. (2006). An ethnobotanical survey of wild edible plants of Paphos and Larnaca countryside of Cyprus. Journal of Ethnobiology and Ethnomedicine 2:34.
Dennis RLH. (1992). The Ecology of Butterflies in Britain. Oxford University Press.
Densmore F. (1928). Uses of plants by the Chippewa Indians. 44th Annual Report of the Bureau of American Ethnology, 275–397.
Dioscorides (1st c. CE). De Materia Medica. Beck trans. (2005), Olms-Weidmann.
Dreyer J, Müssig J. (2000s). Faserinstitut Bremen publications on nettle fibre processing.
Drucker P. (1951). The Northern and Central Nootkan Tribes. Bureau of American Ethnology Bulletin 144.
Edom G. (2005). Stinging Nettles for Textiles. De Montfort University thesis / monograph.
Ellenberg H. (1988). Vegetation Ecology of Central Europe, English ed. Cambridge University Press.
Emmelin N, Feldberg W. (1947). The mechanism of the sting of the common nettle (Urtica urens). Journal of Physiology 106(4):440–455.
ESCOP (European Scientific Cooperative on Phytotherapy). (2003). ESCOP Monographs: The Scientific Foundation for Herbal Medicinal Products. Thieme.
European Medicines Agency / HMPC. (2008). Community Herbal Monograph on Urtica dioica L. and U. urens L., herba. EMA/HMPC/.
European Medicines Agency / HMPC. (2012). Community Herbal Monograph on Urtica dioica L., U. urens L., their hybrids or their mixtures, radix. EMA/HMPC/461160/2008.
European Medicines Agency / HMPC. (2017). Periodic review and associated folium documentation. EMA/HMPC/508013/2006.
European Commission. (2017). Implementing Regulation (EU) 2017/419 approving Urtica extract as a basic substance under Reg. (EC) 1107/2009.
Farag MA, Weigend M, Luebert F, et al. (2013). Phytochemical, phylogenetic, and anti-inflammatory evaluation of 43 Urtica accessions based on UPLC-Q-ToF-MS metabolomic profiles. Phytochemistry 96:170–183.
Felter HW, Lloyd JU. (1898). King’s American Dispensatory, 19th ed. Ohio Valley Company, Cincinnati.
Feliks J. (1981). The Plant World of the Bible. Israel Museum.
Flora of China Editorial Committee. (2003). Urticaceae. Flora of China, Vol. 5.
Font Quer P. (1962). Plantas medicinales: el Dioscórides renovado. Labor.
Fox-Davies AC. (1909). A Complete Guide to Heraldry. T.C. & E.C. Jack.
Fu HY, Chen SJ, Chen RF, et al. (2006). Identification of oxalic acid and tartaric acid as major persistent pain-inducing toxins in the stinging hairs of the nettle Urtica thunbergiana. Annals of Botany 98:57–65.
Fuchs L. (1542). De Historia Stirpium. Basel.
Ganßer D, Spiteller G. (1995). Plant constituents interfering with human sex hormone-binding globulin: evaluation of a test method and its application to Urtica dioica root extracts. Zeitschrift für Naturforschung C 50(1–2):98–104.
Gerard J. (1597). The Herball, or Generall Historie of Plantes. John Norton, London.
Ghorbanibirgani A, Khalili A, Zamani L. (2013). The efficacy of stinging nettle (Urtica dioica) in patients with benign prostatic hyperplasia. Iranian Red Crescent Medical Journal 15(1):9–10.
Gilmore MR. (1919). Uses of plants by the Indians of the Missouri River region. 33rd Annual Report of the Bureau of American Ethnology.
Gleason HA, Cronquist A. (1991). Manual of Vascular Plants of Northeastern United States and Adjacent Canada, 2nd ed. NYBG Press.
Grejtovský A, Markušová K, Eliašová A. (2006). The response of nettle (Urtica dioica L.) to soil heavy metal contamination. Plant, Soil and Environment 52:61–66.
Green MH (ed./trans.). (2001). The Trotula: A Medieval Compendium of Women’s Medicine. University of Pennsylvania Press.
Grela ER et al. (2019). Nettle inclusion in pig and poultry diets: various Polish trial reports.
Grieve M. (1931). A Modern Herbal. Jonathan Cape, London.
Grime JP, Hodgson JG, Hunt R. (2007). Comparative Plant Ecology: A Functional Approach to Common British Species, 2nd ed. Castlepoint Press.
GRIN-Global / USDA-ARS. (2024). Urtica dioica L. Taxonomy record.
Guil-Guerrero JL, Rebolloso-Fuentes MM, Isasa MET. (2003). Fatty acids and carotenoids from stinging nettle (Urtica dioica L.). Journal of Food Composition and Analysis 16:111–119.
Gülçin I, Küfrevioğlu OI, Oktay M, Büyükokuroğlu ME. (2004). Antioxidant, antimicrobial, antiulcer and analgesic activities of nettle (Urtica dioica L.). Journal of Ethnopharmacology 90(2–3):205–215.
Gunther E. (1945/1973). Ethnobotany of Western Washington. University of Washington Publications in Anthropology.
Hamaker J, Weaver D. (1982). The Survival of Civilization. Hamaker-Weaver Publishers.
Hamel PB, Chiltoskey MU. (1975). Cherokee Plants and their Uses: A 400 Year History. Herald Publishing, Sylva NC.
Harley JL, Harley EL. (1987). A check-list of mycorrhiza in the British flora. New Phytologist 105 (Suppl.):1–102.
Heldreich T von. (1862). Die Nutzpflanzen Griechenlands. Athens.
Hellson JC. (1974). Ethnobotany of the Blackfoot Indians. Canadian Ethnology Service, Mercury Series 19.
Herrick JW. (1977). Iroquois Medical Botany. PhD dissertation, SUNY Albany.
Hildegard of Bingen (c. 1150). Physica. Throop trans. (1998), Healing Arts Press.
Hirano T, Homma M, Oka K. (1994). Effects of stinging nettle root extracts and their steroidal components on the Na+/K+-ATPase of the benign prostatic hyperplasia. Planta Medica 60:30–33.
Hoffmann D. (2003). Medical Herbalism: The Science and Practice of Herbal Medicine. Healing Arts Press.
Høeg OA. (1974). Planter og Tradisjon: Floraen i levende tale og tradisjon i Norge 1925–1973. Universitetsforlaget.
Huffman MA. (1997). Current evidence for self-medication in primates. Yearbook of Physical Anthropology 40:171–200.
Humphries DJ, Reynolds CK et al. (UK). Various pasture and silage research outputs on nettle as forage.
Ibn al-Bayṭār (13th c.). Al-Jāmiʿ li-mufradāt al-adwiya wa-l-aghdhiya.
Ibn Sīnā (Avicenna) (c. 1025). Al-Qānūn fī al-Ṭibb. Gruner trans. (1930); Bakhtiar ed. (1999).
Jacomet S. (2006). Plant economy and village life in Neolithic lake dwellings. Vegetation History and Archaeobotany 15:169–182.
Kanter M, Coskun O, Budancamanak M. (2005). Hepatoprotective effects of Nigella sativa L. and Urtica dioica L. World Journal of Gastroenterology 11(42):6684–6688.
Kara K. (2016). Chemical composition, in vitro digestibility and gas production of stinging nettle (Urtica dioica). Turkish Journal of Veterinary and Animal Sciences 40:546–553.
Katz SE. (2012). The Art of Fermentation. Chelsea Green.
Kianbakht S, Khalighi-Sigaroodi F, Dabaghian FH. (2013). Improved glycemic control in patients with advanced type 2 diabetes mellitus taking Urtica dioica leaf extract. Clinical Laboratory 59(9–10):1071–1076.
Kirtikar KR, Basu BD. (1918, 2nd ed. 1935). Indian Medicinal Plants. Allahabad.
Kletter C, Kriechbaum M (eds.). (2001). Tibetan Medicinal Plants. Medpharm Scientific Publishers.
Koepf HH. (1989). The Biodynamic Farm. Floris Books.
Kourik R. (1986). Designing and Maintaining Your Edible Landscape Naturally. Metamorphic Press.
Kregiel D, Pawlikowska E, Antolak H. (2018). Urtica spp.: Ordinary Plants with Extraordinary Properties. Molecules 23(7):1664. doi:10.3390/molecules23071664.
Kuhnlein HV, Turner NJ. (1991). Traditional Plant Foods of Canadian Indigenous Peoples: Nutrition, Botany and Use. Gordon and Breach.
Kumaki Y, Wandersee MK, Smith AJ, et al. (2011). Inhibition of SARS-CoV replication in a lethal BALB/c mouse model by Urtica dioica agglutinin. Antiviral Research 90(1):22–32.
Kwiatkowska E et al. (2015). Ensiling Urtica dioica with grass. Polish Journal of Agronomy (specific issue 2015).
Legssyer A, Ziyyat A, Mekhfi H, et al. (2002). Cardiovascular effects of Urtica dioica L. in the rat. Phytotherapy Research 16(6):503–507.
Leighton AL. (1985). Wild Plant Use by the Woods Cree (Nihithawak) of East-Central Saskatchewan. Canadian Ethnology Service, Mercury Series 101.
Li Shizhen. (1596). Bencao Gangmu 本草綱目.
Ligaa U. (1996). Medicinal Plants of Mongolia. KCA Press.
Linnaeus C. (1753). Species Plantarum. Stockholm.
Loetscher Y, Kreuzer M, Messikommer RE. (2013). Utility of nettle (Urtica dioica) in layer diets as a natural yellow colorant for egg yolk. Animal Feed Science and Technology 186:158–168.
Lopatkin N, Sivkov A, Walther C, et al. (2005). Long-term efficacy and safety of a combination of sabal and urtica extract for lower urinary tract symptoms. World Journal of Urology 23(2):139–146.
Mabey R. (1972, rev. 2012). Food for Free. Collins.
Manandhar NP. (2002). Plants and People of Nepal. Timber Press.
Marzell H. (1943–1979). Wörterbuch der deutschen Pflanzennamen. S. Hirzel.
McIntyre A. (2010). The Complete Herbal Tutor. Gaia.
Memorial Sloan Kettering Cancer Center. About Herbs, Botanicals & Other Products: Nettle. https://www.mskcc.org/cancer-care/integrative-medicine/herbs/nettle.
Mittman P. (1990). Randomized, double-blind study of freeze-dried Urtica dioica in the treatment of allergic rhinitis. Planta Medica 56(1):44–47.
Moerman DE. (1998). Native American Ethnobotany. Timber Press. Database: naeb.brit.org.
Moerman DE. (2010). Native American Food Plants: An Ethnobotanical Dictionary. Timber Press.
Moore M. (1993). Medicinal Plants of the Pacific West. Red Crane Books.
Moszyński K. (1929–39). Kultura ludowa Słowian. PAU.
Nadkarni KM. (1908, rev. 1954). Indian Materia Medica. Popular Prakashan.
Obertreis B, Giller K, Teucher T, Behnke B, Schmitz H. (1996). Anti-inflammatory effect of Urtica dioica folia extract in comparison to caffeic malic acid. Arzneimittel-Forschung 46(1):52–56.
Oliver F, Amon EU, Breathnach A, et al. (1991). Contact urticaria due to the common stinging nettle (Urtica dioica). Clinical and Experimental Dermatology 16(1):1–7.
Opie I, Opie P. (1959). The Lore and Language of Schoolchildren. Oxford University Press.
Orčić D, Francišković M, Bekvalac K, et al. (2014). Quantitative determination of plant phenolics in Urtica dioica extracts by HPLC-MS/MS. Food Chemistry 143:48–53.
Otles S, Yalcin B. (2012). Phenolic compounds analysis of root, stalk, and leaves of nettle. The Scientific World Journal 2012:564367.
Parkinson J. (1640). Theatrum Botanicum. Thomas Cotes, London.
Pasang Yonten Arya. (1998). Dictionary of Tibetan Materia Medica. Motilal Banarsidass.
Penso G. (1983). Index Plantarum Medicinalium. OEMF.
Peumans WJ, De Ley M, Broekaert WF. (1984). An unusual lectin from stinging nettle (Urtica dioica) rhizomes. FEBS Letters 177(1):99–103.
Pettit E. (2001). Anglo-Saxon Remedies, Charms, and Prayers from British Library MS Harley 585: The Lacnunga. Edwin Mellen Press.
Pfeiffer E. (1938, repr. 1983). Bio-Dynamic Farming and Gardening. Anthroposophic Press.
Phillips M. (2011). The Holistic Orchard: Tree Fruits and Berries the Biological Way. Chelsea Green.
Pigott CD, Taylor K. (1964). The distribution of some woodland herbs in relation to the supply of nitrogen and phosphorus in the soil. Journal of Ecology 52 (Suppl.):175–185.
Pinelli P, Ieri F, Vignolini P, et al. (2008). Extraction and HPLC analysis of phenolic compounds in leaves, stalks, and textile fibers of Urtica dioica L. Journal of Agricultural and Food Chemistry 56(19):9127–9132.
Pitcairn CER, Leith ID, Sheppard LJ, et al. (1998). The relationship between nitrogen deposition, species composition and foliar nitrogen concentrations in woodland flora. Environmental Pollution 102(S1):41–48.
Plants of the World Online (POWO). (2026). Royal Botanic Gardens, Kew.
https://powo.science.kew.org/
Pliny the Elder (1st c. CE). Naturalis Historia. Book XXII, chs. 13–17.
Pollard E. (1979). Population ecology and change in range of the white admiral butterfly Ladoga camilla in England. Ecological Entomology 4:61–74.
Pollard E, Hooper MD, Moore NW. (1974). Hedges. Collins New Naturalist.
Pullin AS. (1987). Changes in leaf quality following clipping and regrowth of Urtica dioica. Oikos 49:39–45.
Pywell RF et al. (2010). UK CEH pasture reversion trials (various reports).
Randall C, Randall H, Dobbs F, Hutton C, Sanders H. (2000). Randomised controlled trial of nettle sting for treatment of base-of-thumb pain. Journal of the Royal Society of Medicine 93(6):305–309.
Reeve JR et al. (2010). Influence of biodynamic preparations on compost and soil. Washington State University publications.
Riehemann K, Behnke B, Schulze-Osthoff K. (1999). Plant extracts from stinging nettle (Urtica dioica) inhibit NF-κB. FEBS Letters 442(1):89–94.
Rodwell JS (ed.). (1991–2000). British Plant Communities, Vols. 1–5. Cambridge University Press.
Rogers DJ. (1980). Lakota Names and Traditional Uses of Native Plants. Rosebud Educational Society.
Roschek B Jr, Fink RC, McMichael M, Alberte RS. (2009). Nettle extract (Urtica dioica) affects key receptors and enzymes associated with allergic rhinitis. Phytotherapy Research 23(7):920–926.
Rousseau J. (1945). Le folklore botanique de Caughnawaga. Contributions de l’Institut Botanique de l’Université de Montréal 55.
Rutto LK, Xu Y, Ramirez E, Brandt M. (2013). Mineral properties and dietary value of raw and processed stinging nettle. International Journal of Food Science 2013:857120.
Safarinejad MR. (2005). Urtica dioica for treatment of benign prostatic hyperplasia. Journal of Herbal Pharmacotherapy 5(4):1–11.
Saul FA, Rovira P, Boulot G, et al. (2000). Crystal structure of Urtica dioica agglutinin. Structure 8(6):593–603.
Schöttner M, Ganßer D, Spiteller G. (1997). Lignans from the roots of Urtica dioica and their metabolites bind to human sex hormone binding globulin. Planta Medica 63(6):529–532.
Schneider T, Rübben H. (2004). Stinging nettle root extract (Bazoton-uno) in long-term treatment of benign prostatic syndrome. Der Urologe A 43(3):302–306.
Scott JA. (1986). The Butterflies of North America. Stanford University Press.
Smith HH. (1923). Ethnobotany of the Menomini. Bulletin of the Public Museum of the City of Milwaukee 4(1):1–174.
Smith HH. (1928). Materia Medica of the Bella Coola and neighboring tribes of British Columbia. National Museum of Canada Bulletin 56:47–68.
Smith HH. (1932). Ethnobotany of the Ojibwe Indians. Bulletin of the Public Museum of the City of Milwaukee 4(3):327–525.
Smith HH. (1933). Ethnobotany of the Forest Potawatomi Indians. Bulletin of the Public Museum of the City of Milwaukee 7(1):1–230.
Srutek M, Teckelmann M. (1998). Review of biology and ecology of Urtica dioica. Preslia 70:1–19.
Steiner R. (1924). Agriculture Course (Koberwitz lectures). Rudolf Steiner Press.
STING Project (2002–2005). Sustainable Technologies in Nettle Growing. EU FP5 project, coordinator De Montfort University. CORDIS records ID QLK5-CT-2002-01401.
Tahri A, Yamani S, Legssyer A, et al. (2000). Acute diuretic, natriuretic and hypotensive effects of a continuous perfusion of aqueous extract of Urtica dioica in the rat. Journal of Ethnopharmacology 73(1–2):95–100.
Taylor K. (2009). Biological Flora of the British Isles: Urtica dioica L. Journal of Ecology 97(6):1436–1458. doi:10.1111/j.1365-2745.2009.01575.x.
Thomson S. (1822). New Guide to Health; or, Botanic Family Physician. Boston.
Thurston EL. (1974). Morphology, fine structure, and ontogeny of the stinging emergence of Urtica dioica. American Journal of Botany 61(8):809–817.
Thurston EL, Lersten NR. (1969). The morphology and toxicology of plant stinging hairs. Botanical Review 35:393–412.
Tsangnyön Heruka (15th c.). Life of Milarepa. Quintman trans. (2010), Penguin Classics.
Tutin TG, Heywood VH, et al. (1964–1980). Flora Europaea. Cambridge University Press.
Turner NJ. (1973). The ethnobotany of the Bella Coola Indians of British Columbia. Syesis 6:193–220.
Turner NJ. (1995). Food Plants of Coastal First Peoples. UBC Press.
Turner NJ. (2014). Ancient Pathways, Ancestral Knowledge: Ethnobotany and Ecological Wisdom of Indigenous Peoples of Northwestern North America. McGill-Queen’s University Press.
Turner NJ, Bell MA. (1971). The ethnobotany of the Coast Salish Indians of Vancouver Island. Economic Botany 25:63–104.
Turner NJ, Bell MA. (1973). The ethnobotany of the southern Kwakiutl Indians of British Columbia. Economic Botany 27:257–310.
Turner NJ, Bouchard R, Kennedy DID. (1980). Ethnobotany of the Okanagan-Colville Indians of British Columbia and Washington. Royal British Columbia Museum Occasional Paper 21.
Turner NJ, Efrat BS. (1982). Ethnobotany of the Hesquiat Indians of Vancouver Island. British Columbia Provincial Museum Cultural Recovery Paper 2.
Turner NJ, Thompson LC, Thompson MT, York AZ. (1990). Thompson Ethnobotany: Knowledge and Usage of Plants by the Thompson Indians of British Columbia. Royal British Columbia Museum Memoir 3.
Upton R (ed.). (2009). American Herbal Pharmacopoeia: Stinging Nettle Herb and Root. American Herbal Pharmacopoeia.
Upton R. (2013). Stinging nettles leaf (Urtica dioica L.): Extraordinary vegetable medicine. Journal of Herbal Medicine 3(1):9–38.
USDA PLANTS Database. (2024). Urtica dioica L. and U. gracilis Aiton.
https://plants.usda.gov/
Vogl CR, Hartl A. (2003). Production and processing of organically grown fiber nettle and its potential use in the natural textile industry. American Journal of Alternative Agriculture 18:119–128.
Voisin A. (1959, repr. 1988). Grass Productivity. Island Press.
von Wistinghausen C, Scheibe W, König UJ. (2000). The Biodynamic Spray and Compost Preparations: Production Methods. Floris Books.
Wagner H, Willer F, Samtleben R, Boos G. (1994). Search for the antiprostatic principle of stinging nettle (Urtica dioica) roots. Phytomedicine 1(3):213–224.
Wang B, Qiu Y-L. (2006). Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 16:299–363.
Warrier PK, Nambiar VPK, Ramankutty C (eds.). (1994–96). Indian Medicinal Plants: A Compendium of 500 Species. Arya Vaidya Sala Kottakkal.
Watt G. (1890). A Dictionary of the Economic Products of India. Cosmo.
Weed SS. (1989). Healing Wise. Ash Tree Publishing.
Weiss RF. (1960, Eng. 1988). Lehrbuch der Phytotherapie / Herbal Medicine. Beaconsfield Publishers.
Wood M. (2008). The Earthwise Herbal: A Complete Guide to Old World Medicinal Plants. North Atlantic Books.
WSSA International Herbicide-Resistant Weed Database (Heap I). https://www.weedscience.org
Wyman LC, Harris SK. (1941). Navajo Indian Medical Ethnobotany. University of New Mexico Bulletin 366.
21.5 Living document notes
Date of last research pass: 2026-04-21.
Sections flagged for deeper investigation:
19.6 — North American U. gracilis phytochemistry is the highest-priority research frontier.
19.11 — UDA lectin antiviral frontier, particularly SARS-CoV-2 specific studies and broader high-mannose-virus panels.
19.13 — Pregnancy safety clinical observational studies.
19.3 — Dynamic-accumulator claim; needs formal refutation or validation, not continued folk repetition.
New studies to incorporate when available:
Post-2024 phytochemistry and clinical trial updates (PubMed alerts on “Urtica dioica”).
Post-split U. gracilis taxonomic synthesis papers.
Contemporary Indigenous North American ethnobotanical publications from the communities named in §10.
Updates to EMA HMPC Urticae monographs.
Updates to WSSA Heap database.
Questions raised that have not been answered:
The six cross-cultural convergences identified in §11.6 each generate frontier hypotheses in §19.10. Five of the six are not yet formally tested at modern clinical or mechanistic standards.
The UDA lectin’s ecological function in the plant’s own life — why does Urtica synthesize a broad-spectrum antiviral lectin in its rhizome? — is not clearly understood.
Whether U. gracilis represents a distinct chemotype from U. dioica is genuinely unknown.
Whether biodynamic preparation 504 has an isolated measurable effect on compost biology is empirically contested; the question remains open.
The persistent conflation of Urtica nettle fibre with Boehmeria nivea ramie in historical and ethnographic literature remains partially unresolved; each specific claim requires microscopy verification.
The dock-leaf mechanism for sting relief remains anecdotally universal and mechanistically under-studied.
Thank you so much! Just yesterday I took out a paid-for-one-month-only trial. This morning, I hit the jack-pot. Reading, followed the clue you laid to "purin d'ortie" and transcribed a whopping lot of info from France-Serres (also showing the benefit of having other languages, as this site provided info I have not seen in English). In May I made this year's ferment of nettle, comfrey and garlic cloves. In 3 days the 6 gallon pail's lid was pushed up about 2", the next day, 4". I have been making ferments and adding them to my foliar mix, but at a much greater dilution (1 tb /gallon rain water)--guess I can increase the proportion. I also have been spraying vegetables and fruit trees with a mix of molasses, nettle-comfrey ferment, seaweed ferment (I have access to ascophyllum nodosum and fuscus vesiculosus--which in weed-form I also laid at base of newly-planted cherries and plums), soured raw milk, roasted eggshell/shrimp shell + vinegar, same with bone, now adding fish hydrolysate. Soil is given urine. Haven't used anything purchased (besides the molasses and fish) for about four years. Vegetables, fruit trees are so happy! Acquired Jay McCayman's book for my birthday last summer, so I do indeed see "weeds" as information. And I ferment them . Post scriptum--my nettle patch I created, thirty-ish years ago, and had to try more than one season to get it to take. Used to make champ (stelk) in earliest spring for my family--a treat.
Amazing work! Thank you!