
Dandelion — The Plant Older Than the Literature
Six unrelated languages caught dandelion's diuresis without instruments. Modern clinical research still hasn't run the trial that would honour what they knew.

Dandelion, Taraxacum officinale.
Table of Contents
Phase I, The Plant in Its World
Plant Identity Snapshot
Names, Language, and Lineage
Identification and Look-Alikes
Botanical Character and Life Cycle
Ecological Intelligence
Animal Interactions and Ethology
Climate Resilience and Adaptation
Phenology and Working Calendar
History, Folklore, and Cultural Memory
Traditional Ecological Knowledge and Stewardship
Phase II, The Plant in Human and Animal Hands
Food, Medicine, and Human Use Traditions
Chemistry, Nutrition, and Functional Compounds
Safety and Responsible Use
Regenerative Agriculture and Land Applications
Homestead and Material Uses
Harvest, Processing, and Preservation
Economics and Practical Value
Legal, Regulatory, and Access Notes
Phase III, The Honest Edges
Research Frontiers and Open Questions
Speculative, Symbolic, and Relational Layer
Sources, Confidence, and Citation Architecture
How to read this document
This is a working monograph, not a finished one. Read it in three passes if it serves you:
First pass, the plant in its world (Phase I). Ecology before human use. Names, identification, life cycle, soil, water, animals, climate, season. Understand what the dandelion is doing on its own ground before interpreting what it does in human hands.
Second pass, the plant in human and animal hands (Phase II). Cuisine, traditional medicine across cultures, chemistry, safety, regenerative agriculture, harvest with quality-by-sense, economics, legal frame. The cross-cultural convergence finding sits in section11.6.
Third pass, the honest edges (Phase III). Where the chemistry has not yet caught up to what tradition observed; where the speculative-symbolic-relational layer lives with M / B / FH labels; where the bibliography stands behind the work.
The plant is older than the literature. The literature is older than this document. This monograph is a snapshot. The work continues.
A note before we go further
This newsletter is the work of a farmer, writer, and land-reader, not a doctor, herbalist-in-clinical-practice, or licensed professional of any stripe. What you’ll find here is research, observation, traditional knowledge, and honest gap-flagging — offered for thinking with, not as medical, legal, or financial advice.
If you’re considering using a plant medicinally, especially if you’re pregnant, nursing, taking pharmaceuticals, or managing a serious health condition, talk to someone qualified to know your specific situation. Plants interact with bodies and with drugs in ways no general-audience essay can anticipate.
If you’re going to forage, identify with three independent sources before you eat anything, and never harvest from roadsides, sprayed lawns, or contaminated ground. The plants don’t know they’re growing in lead.
I cite my sources and flag what’s well-documented versus what’s traditional, emerging, or speculative. Read accordingly. Disagreement, correction, and better evidence are welcome — that’s how the work continues.
Dandelion, Taraxacum officinale F.H. Wigg.
A Regenerative Plant Ontology, Phase I: The Plant in Its World

1. Plant Identity Snapshot
Common names: dandelion, common dandelion, lion’s tooth, blowball, piss-a-bed, priest’s crown, fairy clock. Latin binomial: Taraxacum officinale F.H. Wigg. Family: Asteraceae, tribe Cichorieae. Type publication: Wiggers, Primitiae Florae Holsaticae, 1780, p. 56 [Wiggers 1780]. Synonymy / status: Taraxacum officinale is a name most modern taxonomists treat as an aggregate, T. officinale agg., covering somewhere on the order of 200 apomictic microspecies in section Taraxacum [POWO 2026; Richards 1973]. The original Wiggers type is in fact a Lapland microspecies, not the dooryard plant of every continent, a small irony worth keeping in mind every time one writes “officinale.” [Well-documented] Plant type: herbaceous taprooted perennial; rosette-forming. Commonly misapplied names: “false dandelion” and “flatweed” are widely applied to Hypochaeris radicata, which is a separate Asteraceae genus mistaken for dandelion in lawns; Leontodon species and Crepis species are occasionally sold or labeled as “dandelion greens” in market and foraging contexts; the apomictic complex blurs the line between T. officinale sensu lato and T. erythrospermum (red-seeded dandelion) where field workers conflate the two. Native range: Eurasia (Europe and western Asia). Introduced range: every continent except Antarctica’s interior; established in maritime Antarctica [Molina-Montenegro et al. 2012]. Present in all 50 U.S. states and every Canadian province and territory [USDA PLANTS 2026]. Status: introduced, naturalized, ubiquitous; not federally listed as a noxious weed in the United States or Canada despite reputation [USDA APHIS 2026]; treated by EPPO as non-regulated [EPPO 2026].
One-sentence thesis. Dandelion is a triploid clone that travels on a wind, taps the disturbed ground beneath every human foot, and feeds, pollinator, livestock, fungus, child, at every level of a system humans tried to simplify.
Relationship thesis. What humans call a weed, the soil calls a wound dressing, and the early bee calls breakfast.
2. Names, Language, and Lineage
2.1 Scientific identity
The genus Taraxacum G.H. Weber ex F.H. Wigg. was conserved at the Linnaean type Leontodon taraxacum L. and re-typified by Kirschner & Štěpánek [Kirschner & Štěpánek 2011]. The species name officinale is Linnaeus’s catalog tag for canonical drug plants, “of the officina,” the apothecary’s workshop. The accepted name in POWO is Taraxacum officinale F.H. Wigg., treated as an aggregate of microspecies derived from the apomictic complex first synthesized by Richards [Richards 1973; van Dijk 2003; POWO 2026].
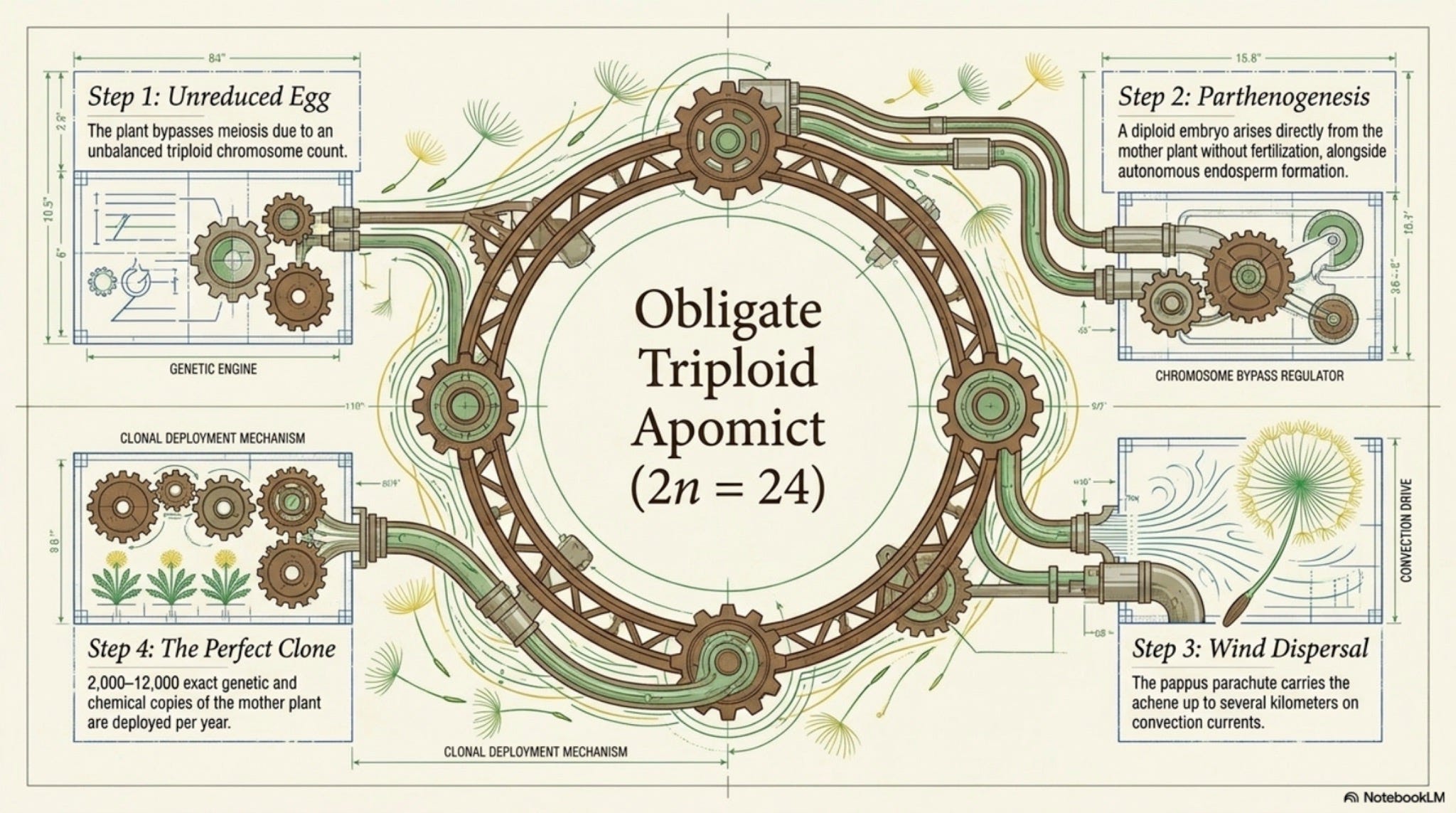
Most weedy T. officinale worldwide are obligate triploid (2n = 24) apomicts, they set seed without fertilization, by autonomous endosperm and parthenogenesis [Richards 1973; van Dijk 2003]. Sexual diploids (2n = 16) survive in southern European refugia [Verduijn et al. 2004]. North American populations are predominantly the European-origin triploid clone [Stewart-Wade et al. 2002]. The “general-purpose genotype” hypothesis explains how a single asexual lineage could colonize so many climates [van Dijk 2003]. [Well-documented]
2.2 Names across cultures
A name is a closed observation. When several unrelated tongues catch the same trait, the trait is real — and across the dandelion’s range, the names cluster around three observations the plant reliably forces on anyone who looks at it.
The Latin binomial Taraxacum officinale names the apothecary’s workshop; officinale is Linnaeus’s catalog tag for canonical drug plants [Wiggers 1780]. Behind the Latin sits an older lineage. The verifiable Arabic source-word for the medieval Latin taraxacon is ṭarakhshaqūn / ṭarakhshaqūq, traceable through al-Rāzī, Avicenna, and Ibn al-Bayṭār [Lev & Amar 2008; Genaust 1996]. Arabic itself carries the plant under hindibā’ barriyya, wild endive, signaling that medieval Arabic medicine treated the chicory-dandelion complex as one tribe. Modern Persian keeps a separate observation: qāsedak, little messenger — the airborne seed that carries word [Ghahreman, Flora of Iran].
In Europe, two parallel name-streams move side by side. The dental stream reads the leaf. Old French dent-de-lion gave English its dandelion; Welsh dant y llew (lion’s tooth) is the earliest vernacular European attestation, traced to the thirteenth-century Meddygon Myddfai [Pughe 1861]. German Löwenzahn, Italian dente di leone, Spanish diente de leónrepeat the image without consultation. Turkish karahindiba, black endive, carries the same toothed-leaf signal under a different metaphor [Baytop 1999].
The seed-clock stream reads the wind. English blowball, fairy clock, telltime; German Pusteblume (blow-flower); Italian soffione; Russian одуванчик (oduvanchik), from dut’, “to blow” [Vasmer]; Persian qāsedak, the messenger. A child’s instrument for telling time, the same instrument in seven languages.
The third stream is the one that carries the strongest signal, and it carries it as a bedwetter warning. English piss-a-bed, French pissenlit, German Bettpisser, Italian piscialletto, Spanish meacamas, Dutch beddezeiker — six unrelated tongues catching the same effect [Marzell 1943–1979; Rolland 1896–1914; Britten & Holland 1886]. The diuresis is real, and folk knowledge knew it without instruments.
Beyond these three clusters, the names get more local. English regional usage adds priest’s crown, monk’s head, swine’s snout, peasant’s clock, cankerwort, Irish daisy — the bald receptacle after seed dispersal naming the plant for its monastic profile. French keeps couronne de moine (monk’s crown) and salade de taupe (mole’s salad). German adds Kuhblume(cow-flower), Pfaffenröhrlein (priest’s-little-pipe, for the hollow scape), and Maiblume (May-flower) — the cattle-turnout calendar. Dutch paardenbloem (horse-flower) and molsla (mole’s salad). Polish mniszek lekarski, little medicinal monk. Hungarian gyermekláncfű, child’s-chain grass — the daisy-chain made from scapes.
In East Asia the picture shifts in ways worth carrying carefully. The Chinese name 蒲公英 Pugongying (Pugong’s flower) appears in the Tang Bencao of 659 CE and Li Shizhen’s Bencao Gangmu of 1596 [Tang Bencao 659; Li Shizhen 1596; PRC Pharmacopoeia 2020]. But the official drug taxon under that name in the modern PRC pharmacopoeia is T. mongolicum, with T. officinale an accepted equivalent — a distinction that matters and one this profile keeps flagging. Japanese タンポポ tanpopo is onomatopoeic; the introduced T. officinale arrived around 1900 and is specified as セイヨウタンポポ seiyō-tanpopo [Morita et al. 1985]. Korean 민들레 mindeulle sits in the sansai mountain-vegetable category.
And then there are the silences. Sanskrit and the classical Ayurvedic canon have no record of dandelion — no Charaka, no Sushruta, no Bhāvaprakāśa Nighaṇṭu entry. The Greek and Proto-Indo-European root is silent at the indigenous level; the proposed tarassein + akos derivation is folk-etymological [Genaust 1996]. The pre-colonial African record is silent. The pre-colonial South American record is silent. No major alchemical tradition treats dandelion specifically. The Indigenous North American record is rich but entirely post-Columbian — every Moerman entry is for the introduced T. officinale[Moerman 1998]. These silences are information, not omissions, and the document carries them as gaps rather than papering them over.
2.3 What the names notice
Three patterns repeat across unrelated languages.
The bedwetter cluster, piss-a-bed in English, pissenlit in French, Bettpisser in German, piscialletto in Italian, meacamas in Spanish, beddezeiker in Dutch, is the strongest convergent-naming signal in the Asteraceae [Marzell IV: 624; Rolland III; Britten & Holland 1886]. Six unrelated tongues catch the same effect. The diuresis is real, and folk knowledge knew it without instruments.
The dental cluster, dent-de-lion, dant y llew, diente de león, Löwenzahn, karahindiba (black endive in Turkish, but the leaf shape carries the same tooth-image), reads the lobed leaf. Across half a continent, the leaf says “lion’s tooth” without consultation.
The seed-clock cluster, blowball, Pusteblume, soffione, qāsedak, oduvanchik, fairy clock, telltime, names the achene head and the wind. A child’s instrument for telling time, the same in seven languages.
What the names do not know: any dandelion at all, in classical Sanskrit or in pre-contact North America. [Gap, explicit]

3. Identification and Look-Alikes
3.1 Field identification
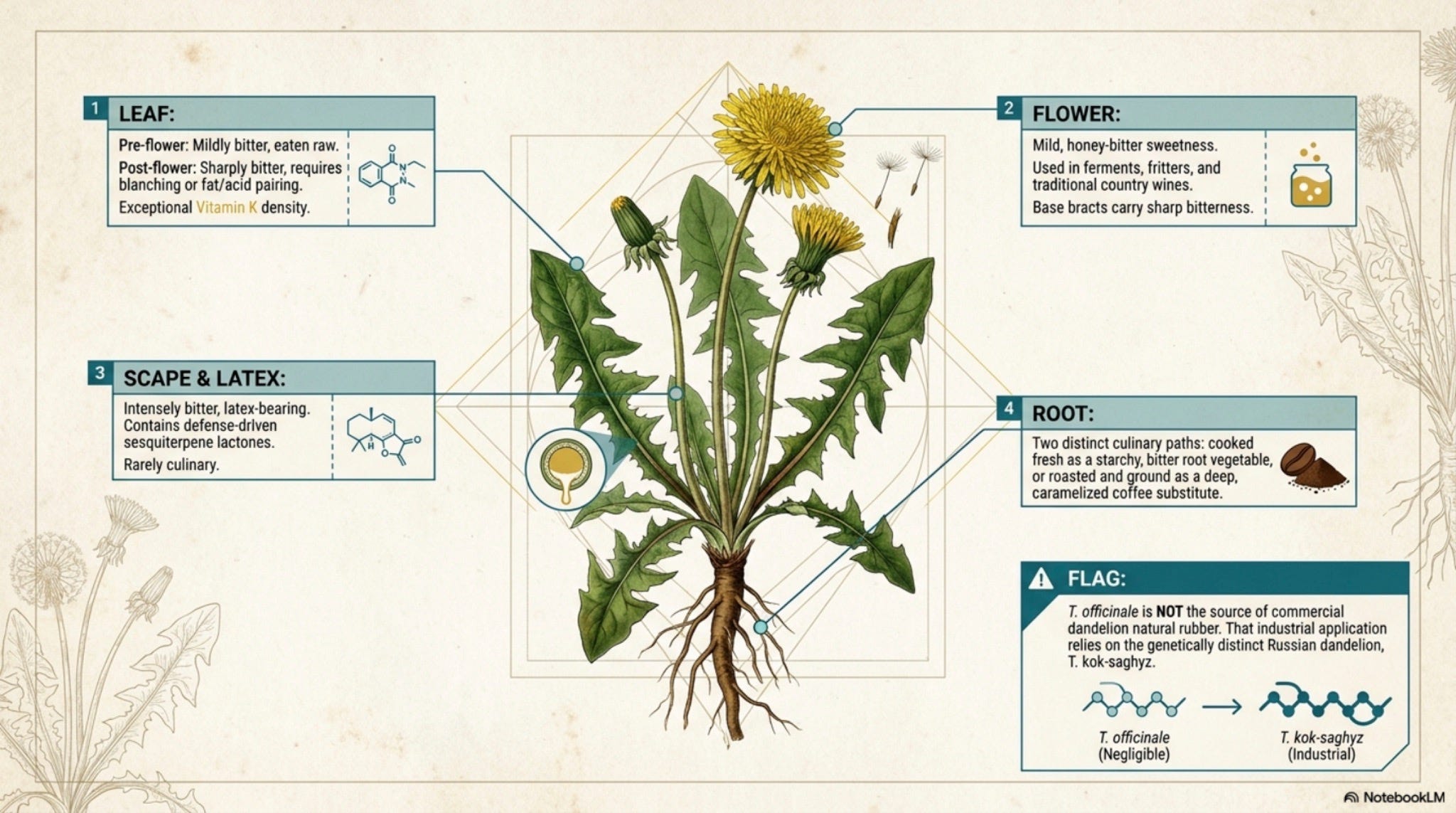
A basal rosette of leaves, flat to the ground; deeply pinnatifid lobes pointing back toward the base, the lion-tooth profile the names notice [FNA Vol. 19 2006]. A single hollow scape, leafless, smooth, milky when broken, rises from the rosette crown to lift one capitulum. All florets are ligulate (ray-only); there are no disc florets. The capitulum is yellow at full bloom, closes at night and in rain, and in fruit dries to the bald white receptacle that gave the plant its monastic and royal names [Penn State Extension 2024]. Each achene is olive-brown to straw, ribbed, beaked, crowned with a parachute of capillary bristles, the pappus that is also a sail.
The taproot is the structure most people never see. Vertical, fleshy, bitter, often forked with depth, latex-bearing throughout, reported in most populations to 0.6–1.5 m, occasionally deeper [Kutschera & Lichtenegger 1960–1992; Cyr et al. 1990]. A fragment 1–2 cm long can regenerate the whole plant [Cyr et al. 1990]. [Well-documented]
Crush a leaf, the smell is faint, green, slightly bitter, almost lactuca-like. A drop of latex on the skin first feels cool, then sticky as it dries to a brown film. This is the body’s first instrument and will not lie to you.
3.2 Look-alikes
The yellow-rayed Asteraceae of disturbed ground are a small crowd, and several have been mistaken for dandelion in the field [Uva, Neal & DiTomaso 1997; DiTomaso & Healy 2007].
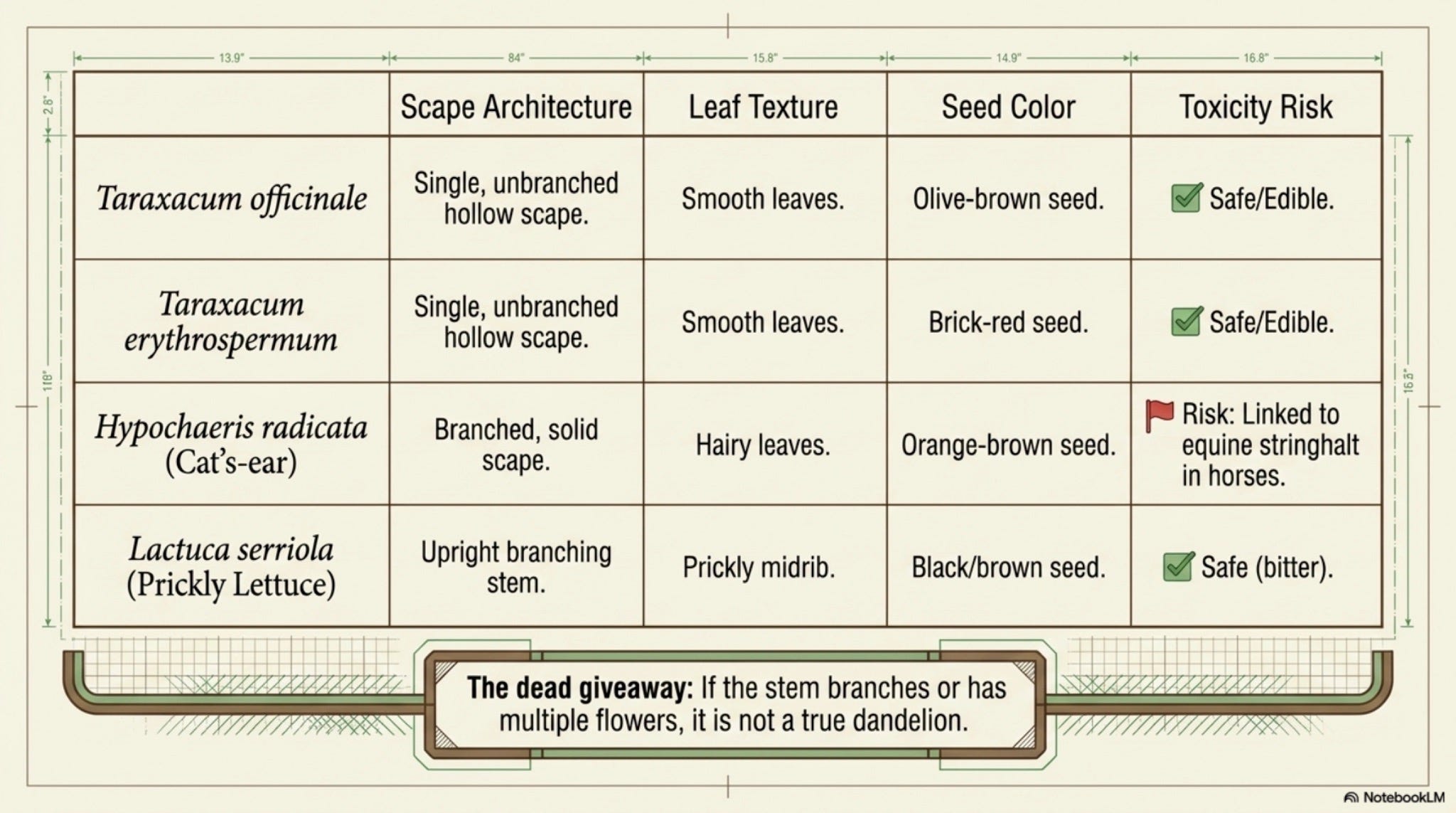
Taraxacum erythrospermum (red-seeded dandelion) is the closest kin; achenes brick-red to purple, leaves more deeply dissected, outer phyllaries appressed-spreading rather than reflexed. Same scape, same latex, same edibility, but the seed color is the diagnostic. [Well-documented]
Hypochaeris radicata (cat’s-ear), leaves hairy where dandelion’s are smooth; scape is solid and branched, holding several heads, not one; rosette stays flatter. Branched scape is the giveaway. [Well-documented]
Crepis spp. (hawksbeards), leafy, branching stems with multiple smaller heads; not a single-scape rosette plant. [Well-documented]
Lactuca serriola (prickly lettuce), tall, leafy, prickly midrib on leaves; latex similar but the architecture is upright, not basal. [Well-documented]
Sonchus spp. (sow-thistles), leafy spiny-margined stems and clasping leaves; latex like dandelion’s but plant is far larger and branched. [Well-documented]
Lapsana communis (nipplewort), branched, leafy, very small heads. [Well-documented]
3.3 Safety note
The principal misidentification risk for foragers is Hypochaeris radicata. The cat’s-ear is edible and used like dandelion in some traditions, but it has been linked to equine stringhalt, a neurological gait disorder in horses, when grazed in quantity in Australia and New Zealand [Cahill et al. 1986]. Dandelion has no comparable record. Beyond that, no member of the look-alike crowd is acutely toxic; the worst outcome of confusion is a less-tasty salad. Latex contact dermatitis from the milky sap is reported but uncommon [Lovell 1993; Mark et al. 1999]. The principal hard rule: do not harvest from roadsides, sprayed lawns, or ground where lead/cadmium is plausible, dandelion roots accumulate trace metals from contaminated soils [Robinson et al. 2009].
4. Botanical Character and Life Cycle
A long-lived herbaceous perennial, surviving from a fleshy taproot that overwinters frozen and resumes activity at the first warm soil. The rosette can re-grow from the root crown after defoliation, mowing, or grazing; if the upper taproot is severed, fragments as short as 1–2 cm can regenerate adventitious shoots [Cyr et al. 1990]. This is why a hoe never finishes the job.
Reproduction is the most interesting part of the plant. The common weedy form is a triploid apomict [Richards 1973]. Triploid means three sets of chromosomes, an unbalanced number that cannot pair properly at meiosis. Apomictic means seed is set without fertilization: a diploid embryo arises from an unreduced egg cell, with autonomous endosperm formation [van Dijk 2003]. Each seed is a clone of the mother. A single plant produces 2,000–12,000 achenes per year [Stewart-Wade et al. 2002], and every one is a chemical and genetic copy of the plant that made it. The pappus carries the achene on the wind for hundreds of meters, occasionally kilometers in convection currents [Honěk & Martinková 2005].
Pollination still occurs at low frequency in some triploid populations, pollen production is variable but functional, which permits gene flow into sexual diploid populations and complicates the “pure clone” picture [van Dijk 2003]. Sexual diploids exist in alpine and southern European refugia and pollinate normally [Verduijn et al. 2004]. In North America, virtually all T. officinale are the introduced triploid lineage [Stewart-Wade et al. 2002]. [Well-documented]
Germination is opportunistic, light-stimulated, no significant chilling requirement, viability declines rapidly so the seed bank is short-lived (mostly less than one year) [Honěk & Martinková 2005]. Disturbance is the cue; bare soil is the invitation. The plant is a textbook ruderal: open ground, rapid colonization, quick to reproduce. In Grime’s CSR scheme it sits at competitive-ruderal [Grime 2001].
Successional role is similarly textbook. Pioneer in disturbed ground; strong presence in early-seral plant communities (tilled soil, lawn, pasture); gradually displaced by stable perennial cover unless disturbance recurs. The repeating mow is the mechanism that builds the lawn dandelion population, selecting, every cut, for plants that can flower below the blade and seed before the next pass.